Upload
juan-jose-gonzalvez
View
258
Download
0
Embed Size (px)
DESCRIPTION
resumen de algunos capítulos del libro Best - Taylor.
Citation preview
RENAL
LQUIDOS CORPORALESY FUNCIN RENAL
Fisiologa de los lquidos corporales
La funcin principal de los riones es el mantenimiento del volumen y la composicin normales de los lquidos corporales. As, los riones son responsables de la excrecin del exceso de agua, iones y productos de desecho, as como de la conservacin de los solutos importantes para la funcin corporal adecuada.
AGUA CORPORAL Y SUS SUBDIVICIONES
El agua es el componente ms abundante del organismo, constituye entre el 45 y el 75% del peso corporal. Esta gran variacin en el contenido de agua es, principalmente, funcin de la variacin de las cantidades de tejido adiposo, el porcentaje de peso corporal constituido por agua variar en forma inversa al contenido de grasa del organismo. El agua corporal total (ACT) constituye aproximadamente el 60% del peso corporal en los varones adultos jvenes normales y 50% del peso corporal en las mujeres adultas jvenes normales, debido a que tienen una cantidad algo superior de grasa subcutnea.Para ambos sexos el porcentaje de peso corporal constituido por agua disminuye con la edad.El ACT se distribuye en dos compartimentos lquidos principales: el lquido intracelular (LIC), que contiene aproximadamente el 55% del ACT, y el lquido extracelular (LEC), que contiene aproximadamente el 45% del ACT.El LEC a su vez, est subdividido en varios compartimentos ms pequeos. Los compartimentos ms importantes del LEC son el plasma, que contiene aproximadamente el 7.5% del ACT, y el lquido intersticial (lquido intercelular y de los linfticos) que contiene aproximadamente el 20% del ACT.Tambin se clasifican como parte de los compartimentos del LEC el agua cristalizada en el hueso y el lquido en los tejidos conectivos densos, cada uno de los cuales contiene aproximadamente 7.5% del ACT. Por ltimo, formando una pequea parte del LEC encontramos al lquido transcelular, denominado as porque los compartimentos menores que lo forman se encuentran separados del resto del LEC por una capa de clulas endoteliales.Este lquido transcelular incluye el lquido del aparato gastrointestinal, biliar y urinario, el lquido intraocular y cefalorraqudeo, el lquido de los espacios serosos, como el de la pleura, del peritoneo y del pericardio, en conjunto estos solo contienen aproximadamente el 2.5% del ACT.Con el objetivo de simplificar esta distribucin, en la prctica clnica comnmente se establece que el LEC contiene aproximadamente la tercera parte del ACT (o 20% del peso corporal de un hombre adulto normal) y el LIC contiene aproximadamente las dos terceras partes del ACT (o 40% del peso corporal de un hombre adulto normal).
MEDIDA DE LOS COMPARTIMENTOS LQUIDOS CORPORALES
Para poder medir los volmenes de los distintos compartimentos lquidos del organismo con fines experimentales y clnicos utilizamos mtodos de dilucin.Estos mtodos utilizan sustancias marcadoras que se distribuyen en el lquido de un compartimento especfico del organismo. Si se administra una cantidad conocida de un marcador X y se da tiempo para que ste se distribuya en todo el compartimento, entonces el volumen del compartimento podr determinarse a partir de la concentracin del marcador en una muestra de lquido de ese compartimento:
Vol. del compartimento = masa de X administrado concentracin de X en el compartimento
Una aplicacin ms precisa del mtodo de dilucin incluye la correccin de la cantidad del marcador que se pierde (por ejemplo en la orina) durante el perodo de distribucin:
Vol. del compartimento = (masa de X administrada masa de X perdida) concentracin de X en el compartimento
Los mtodos de dilucin pueden utilizarse para estimar los volmenes de ACT, LEC y plasma.
AGUA CORPORAL TOTAL (ACT)
El volumen de ACT se estima utilizando marcadores que se distribuyen de manera uniforme en todos los lquidos corporales, como el agua deuterada (D2O) o el agua tritiada (HTO). Tambin puede utilizarse el frmaco antipirina, aunque en ciertas partes del agua corporal penetra lentamente y por lo tanto tiende a subestimar el ACT. Dado que el plasma es parte del ACT, las concentraciones del marcador en el compartimento del ACT pueden obtenerse de una muestra de plasma.
LQUIDO EXTRACELULAR (LEC)
La estimacin del LEC con los mtodos de dilucin requiere marcadores que puedan cruzar libremente el endotelio capilar, pero que se excluyan en forma predominante de las clulas. Pueden utilizarse radioistopos de los iones, como el Na+, Cl-, Br-, SO42- y S2O32- (tiosulfato), pero estos encuentran en la clula en grado variable y por lo tanto tienden a sobreestimar el volumen del LEC. Los sacridos no metabolizables, como inulina, manitol y rafinosa, tambin son tiles, pero no se distribuyen rpidamente en todo el compartimento extracelular y, por lo tanto, tienden a subestimar el volumen del LEC.Como ocurre con el ACT, el LEC incluye plasma, de modo que la concentracin de un marcador en el LEC puede obtenerse a partir de una muestra de plasma.
PLASMA
El volumen del plasma puede ser estimado utilizando radioistopos de la albmina o colorante azul de Evans, que se une fuertemente a la albmina. Un mtodo algo diferente para determinar el volumen plasmtico implica el uso de eritrocitos marcados para determinar el volumen sanguneo por dilucin. El volumen plasmtico puede calcularse entonces de la siguiente manera:
Vol. de plasma = volumen sanguneo (1 Hct)
donde Hct es el hematocrito. El mtodo con eritrocitos marcados subestima levemente el volumen plasmtico, dado que el hematocrito medido en una vena perifrica sobreestima levemente el hematocrito real.
LIC Y LQUIDO INTERSTICIAL
Los mtodos de dilucin no pueden utilizarse para medir los volmenes del LIC y de lquido intersticial, dado que no hay marcadores que se distribuyan exclusivamente en estos compartimentos. Sin embargo, una vez que se han estimado por dilucin los volmenes del ACT y del LEC, el volumen del LIC puede calcularse de la siguiente manera:
LIC = ACT LECEl volumen del lquido intersticial puede calcularse a partir de los volmenes del LEC y del plasma:
Lquido intersticial = LEC - plasma
LQUIDO EXTRACELULAR
PLASMA Y LQUIDO INTERSTICIAL
Estos dos compartimentos principales del LEC tienen composiciones muy similares, el Na+ es el catin predominante y el Cl- y el HCO3- son los aniones principales. Sin embargo, una diferencia importante entre el plasma y el lquido intersticial es la mayor concentracin de protenas en el plasma. Esta diferencia existe porque el plasma es libremente permeable al agua y a los solutos pequeos, tales como los iones inorgnicos, glucosa y urea, pero tiene permeabilidad selectiva para los solutos de mayor tamao, como protenas grandes y lpidos. Dada la alta permeabilidad del endotelio capilar a los solutos pequeos, se podra anticipar que las concentraciones de estos solutos en el plasma y en el lquido intersticial seran idnticas, pero luego de algunas correcciones se observa que las concentraciones de los no electrolitos pequeos en el plasma y lquido intersticial son idnticas, pero los electrolitos pequeos tienen concentraciones ligeramente distintas. Estas diferencias pueden atribuirse primariamente al efecto Gibbs-Donnan.
EFECTO GIBSS-DONNAN
1 2 3
5Na+ 5Na+ 10Na+ 9Na+ 6Na+ A = Plasma 5Cl- 5Cl-5Cl- 5Na+ 4Cl- 6Cl- B = Lquido Pr-5 5Cl- Pr-5 intersticial
A B A B A B Endotelio capilar
Consideremos una situacin hipottica donde tengo un compartimento de plasma y otro de lquido intersticial del mismo tamao, separados por una barrera de endotelio capilar. Inicialmente estn presentes 5 iones Na+ y 5 iones Cl- en cada compartimento (1), y luego una molcula de protena que tiene 5 cargas negativas se aade al compartimento del plasma, junto con 5 iones Na+ adicionales para contrarrestar las cargas negativas de la protena (2). Dado que el endotelio capilar es permeable al Na+, ste difunde desde el plasma al lquido intersticial, a favor de su gradiente de concentracin, acompaado de Cl- para mantener la electroneutralidad. Sin embargo, esta migracin aumentar la concentracin de Cl- en el compartimento del lquido intersticial, generando as un gradiente de concentracin que se opone a la posterior difusin de Cl- del plasma al lquido intersticial. De acuerdo a los principios termodinmicos, en el equilibrio la distribucin de Na+ y Cl- entre el plasma y el lquido intersticial, est dada por la relacin Gibbs-Donnan:
PNa . PCl = LINa . LICl
donde P y LI designan las concentraciones de los iones del plasma y lquido intersticial respectivamente. Si X iones Na+ y X iones Cl- migran del plasma al lquido intersticial, antes de que se haya establecido el equilibrio, entonces en el equilibrio (10 X)(5 X) = (5 + X)(5 + X), de modo tal que X = 1, o sea, en el equilibrio en el plasma habr 9 iones Na+, 4 iones Cl- y la molcula proteica con 5 cargas negativas, mientras que en el lquido intersticial habr 6 iones Na+ y 6 iones Cl-. En otras palabras, debido a la presencia de la protena aninica en el compartimento plasmtico, el estado de equilibrio entre ambos compartimentos se caracteriza por tres rasgos importantes: 1) los pequeos iones difusibles (Na+ y Cl-) no tienen concentraciones iguales en ambos compartimentos, la concentracin de catin (Na+) es ligeramente superior en el compartimento plasmtico que contiene a la protena aninica y la concentracin de anin (Cl-) es ligeramente superior en el compartimento del lquido intersticial; 2) la concentracin total de equivalentes de carga es mayor en el compartimento plasmtico que contiene protenas y 3) a pesar de las diferencias en las concentraciones de los iones y en la concentracin total de equivalentes de carga, la electroneutralidad se mantiene dentro de cada compartimento, es decir, el nmero total de cargas catinicas iguala al nmero total de cargas aninicas.
LQUIDO INTRACELULAR
A diferencia de lo que ocurre con el plasma y el lquido intersticial, el LIC no es una fase lquida continua y su composicin precisa difiere en los distintos tejidos.El LIC tiene concentraciones de Na+, Cl- y HCO3- relativamente bajas. En lugar de ello, el catin predominante es el K+, mientras que los aniones principales son los fosfatos orgnicos (por ejemplo el ATP) y protenas.Estas diferencias entre la composicin del LIC y del LEC pueden atribuirse a varios factores. En primer lugar la bomba Na+/K+ ATPasa de las membranas celulares transporta activamente Na+ desde las clulas y K+ hacia el interior de estas, dando cuenta as de la alta concentracin de Na+ y baja de K+ en el LEC, y de la baja concentracin de Na+ y alta de K+ en el LIC. En segundo lugar, la membrana celular que separa al LIC del lquido intersticial tiene una permeabilidad muy limitada a los fosfatos orgnicos y a las protenas, lo que da como resultado el establecimiento de un equilibrio Gibbs-Donnan a travs de la membrana celular. Como se demostr antes, este equilibrio Gibbs-Donnan da cuenta de tres caractersticas importantes que son propias de las diferencias de composicin entre el LIC y el lquido intersticial: 1) la concentracin de pequeos cationes difusibles (Na+, K+) es mayor en el LIC con protenas, mientras la concentracin de pequeos aniones difusibles (por ejemplo el Cl3-) es mayor en el lquido intersticial (sin embargo debido a la presencia de la bomba Na+/K+ ATPasa la distribucin de cationes no est gobernada solamente por el equilibrio Gibbs-Donnan) 2) la concentracin total de equivalentes de carga es mayor en el LIC que tiene protenas y 3) la electroneutralidad se mantiene en cada compartimento a pesar de las diferencias en las concentraciones inicas y en la concentracin total de los equivalentes de carga.
MEDIDA DEL CONTENIDO TOTAL DE ELECTROLITOS DE LOS LQUIDOS CORPORALES
Con mtodos de dilucin similares a los utilizados para determinar el volumen de lquidos corporales, puede obtenerse informacin importante acerca del contenido total de un in dado en los lquidos corporales. El contenido total de un in, determinado por mtodos de dilucin isotpica, se llama pool intercambiable del in en cuestin.Para el Na+, K+ y Cl-, los principales iones de los lquidos corporales, el pool intercambiable representa la mayora del contenido total de estos iones en el organismo. Por ejemplo, el Na+ intercambiable (Na+e) representa ms del 70% del Na+ total del organismo, mientras que el K+ intercambiable (K+e) representa ms del 90% del K+ total del organismo. La mayora de los pools no intercambiables de Na+ y K+ se encuentran en el hueso. Por lo contrario, un porcentaje relativamente pequeo del contenido corporal total de Ca2+, Mg2+ y fosfato es intercambiable, aproximadamente 1%.
OSMOLALIDAD DE LOSLQUIDOS CORPORALES
A pesar de las diferencias en la concentracin de los diferentes compartimentos, estos lquidos tienen esencialmente osmolalidades totales idnticas (290 mosmol/Kg H2O). Ello se debe a que el endotelio capilar y casi todas las membranas celulares son libremente permeables al agua, permitiendo que el plasma, el lquido intersticial y el LIC sean isoosmticos.
ANLISIS DE LA OSMOLALIDAD PLASMTICAY RELACION DE LA CONCENTRACIN DE Na+PLASMTICO CON LA OSMOLALIDAD
Ya que los principales compartimentos lquidos del organismo son isoosmticos, podemos estudiar sus osmolalidades analizando la osmolalidad del plasma. La osmolalidad plasmtica puede estimarse como la suma de contribuciones de electrolitos, ms las concentraciones de glucosa y urea:
Posm osmolalidad de los electrolitos + osmolalidad de la glucosa + osmolalidad de la urea
CONTRIBUCIN DE LOS ELECTROLITOS
Dado que el Na+ y sus aniones acompaantes representan los principales electrolitos plasmticos, puede suponerse que:
Osmolalidad de los electrolitos osmolalidad de Na+ + iones acompaantes
Si se supone adems que cada in Na+ est apareado con un in univalente, entonces:
Osmolalidad de los electrolitos (mosmol/Kg H2O) 2 PNa (mmol/litro) La suposicin hecha anteriormente introduce errores, debido que al considerar que el Na+ y sus iones acompaantes son los nicos electrolitos tiende a subestimar la contribucin osmtica verdadera de los electrolitos, dado que excluye otros cationes como el K+, Ca2+ y Mg2+. Por otro lado, la suposicin de que cada in Na+ est unido con un anin univalente tiende a sobreestimar la contribucin osmtica verdadera de los electrolitos, dado que el Na+ y sus aniones no estn completamente disociados en los lquidos corporales y parte del Na+ est apareado con aniones polivalentes como por ejemplo el SO24- y protenas. Fortuitamente, luego de corregir los diversos errores, la osmolalidad de la contribucin de electrolitos de hecho se aproxima mucho a 2 PNa.
CONTRIBUCINES DE LAGLUCOSA Y LA UREA
Dado que el peso molecular de la glucosa es de 180, el peso molecular del N ureico es 28 (es decir 28g de los 60g en un mol de urea estn formados por N) y 1 litro = 10 dl, las contribuciones de la glucosa y la urea pueden expresarse como:
Osmolalidad de la glucosa (mosmol/Kg H2O) PG (mg/dl)/18
Osmolalidad de la urea (mosmol/Kg H2O) PUN (mg/dl)/2.8
Sustituyendo estos valores:
Posm (mosmol/Kg H2O) 2 PNa (mmol/litro) + PG (mg/dl)/18 + PUN (mg/dl)/2.8
Dado que las contribuciones de la glucosa y la urea normalmente solo dan cuenta de 10 mosmol/Kg H2O, podemos decir que:
Posm (mosmol/Kg H2O) 2 PNa (mmol/litro) + 10
ANATOMA DE LOS RIONES
ASPECTO MACROSCPICO
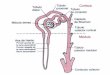
Las funciones excretoras de los riones, ampliamente reguladas, son llevadas a cabo por unidades funcionales individuales, conocidas como nefronas.Cada nefrona consiste en dos estructuras principales, una porcin glomerular, llamada con frecuencia cpsula de Bowman, y un tbulo. Este a su vez puede dividirse en tres regiones principales: el tbulo proximal, el asa de Henle y la nefrona distal. Varias nefronas distales adyacentes comparten un segmento final comn, el tbulo o conducto colector.Pueden distinguirse dos tipos bsicos de nefronas, las corticales y las yuxtamedulares. Aunque todos los glomrulos se localizan en la corteza, las nefronas corticales tienen glomrulos que se encuentran en la regin externa de la corteza, mientras que los glomrulos de las nefronas yuxtamedulares se encuentran en la corteza interna, cerca de la unin corticomedular.Las nefronas corticales tienden a presentar asas de Henle cortas, que permanecen exclusivamente en la corteza o penetran solo en la zona externa de la mdula. Por el contrario las nefronas yuxtamedulares tienden a presentar asas de Henle largas, que descienden profundamente en la zona interna de la mdula, hasta alcanzar en algunas nefronas el extremo de la papila, antes de girar y ascender nuevamente a la corteza. En los humanos el 85% de las nefronas son corticales y el 15% restante son yuxtamedulares.En trminos generales, la funcin de la porcin glomerular de la nefrona es la filtracin de lquidos y sus constituyentes cristaloides desde el plasma al tbulo, mientras que la funcin del tbulo es reducir el volumen y modificar el contenido del filtrado. El tbulo reabsorbe sustancias al lquido tubular, de modo que la orina final contiene los componentes que deben ser excretados para preservar un volumen y composicin normal de los lquidos corporales.
Nefrona Nefrona yuxtamedular cortical
Cpsula de Bowman
Tbulodistal
Tbulopoximal
Tbulo(conducto)colector
Asa deHenle
IRRIGACIN SANGUINEA
La sangre entra al rin por la arteria renal, la cual se ramifica dando origen a las arterias segmentarias, que se subdividen en arterias interlobulares. Cada arteria interlobular penetra en el rin a travs de una columna de Bertin, pero antes de alcanzar la superficie cortical se divide en arterias arcuatas o arciformes, que corren paralelas a la superficie. Estas arterias arcuatas dan a las arterias radiales corticales o arterias interlobulillares y penetran en la corteza de forma perpendicular hacia la superficie. De cada arteria radial cortical se ramifican numerosas arteriolas pequeas, las arteriolas aferentes. Cada arteriola aferente lleva sangre a una nica nefrona. La arteriola aferente interacta en forma ntima con la porcin glomerular de la nefrona, donde se rompe en una red capilar, el penacho capilar glomerular. Aproximadamente 20% del agua del plasma que entra a la arteriola aferente se filtra por el capilar glomerular al rea de acumulacin en forma de copa que se encuentra en el interior de la cpsula de Bowman, llamada con frecuencia espacio de Bowman. El 80% restante del plasma, junto con todos los solutos de mayor tamao y todos los elementos celulares de la sangre, fluye desde los capilares glomerulares a la arteriola eferente (la arteriola eferente sale de los capilares glomerulares). La arteriola eferente se dispersa en una segunda red de capilares que rodea a las porciones tubulares de las nefronas, enviando las sustancias que secretan los tbulos en el lquido tubular y captando agua y solutos reabsorbidos por los tbulos. La mayora de los capilares de esta segunda red rodean a los tbulos de la corteza y se llaman capilares peritubulares. Los capilares que derivan de las arteriolas eferentes de las nefronas yuxtamedulares descienden a profundidad variable en la mdula antes de formar una red capilar. Los capilares vuelven a unirse cuando giran hacia la corteza y ascienden nuevamente hacia ella, estos capilares nicos se conocen como vasa recta (vasos rectos) y tienen una funcin importante en el mecanismo de concentracin de la orina.Desde los capilares peritubulares y los vasa recta, la sangre fluye a las venas radiales corticales y luego abandona el rin en venas que fluyen en sentido contrario a las arterias adyacentes y reciben el nombre de ellas.
NEFRONA
La porcin glomerular de la nefrona, o cpsula de Bowman, est ntimamente relacionado con el penacho de los capilares glomerulares, por esto se utiliza el trmino glomrulo para incluir tanto a la nefrona como al capilar. La barrera de filtracin consta de tres capas: el endotelio capilar, la membrana basal y el epitelio (pedicelos de los podocitos).El endotelio capilar es similar al que tapiza los capilares de cualquier otra zona del cuerpo, excepto en que las clulas del endotelio glomerular tienen numerosas fenestras en su citoplasma. La membrana basal compuesta por colgeno y proteoglucanos (pero no clulas), rodea a los capilares glomerulares.Por encima de la membrana basal se encuentra la capa celular epitelial de la cpsula de Bowman. Las clulas que rodean el penacho capilar difieren de las que forman el resto de la cpsula de Bowman, principalmente porque tienen numerosas prolongaciones pediculadas o pedicelos, de all, que estas clulas se llamen podocitos. Interdispersos entre los capilares adyacentes, particularmente en el centro o ncleo de glomrulo, hay racimos de clulas mesangiales, las cuales proporcionan apoyo a los capilares glomerulares.La porcin tubular de la nefrona consta de una capa celular epitelial, que se contina con la capa celular epitelial que forma la cpsula de Bowman. La membrana basal, continua con la membrana basal glomerular, rodea todo el tbulo. Como sabemos el tbulo puede dividirse en tres regiones principales: tbulo proximal, asa de Henle y nefrona distal. Las clulas epiteliales de varias regiones del tbulo difieren en muchos aspectos, sin embargo, hay ciertos rasgos estructurales comunes a todas las clulas epiteliales tubulares.
TBULO PROXIMAL
El tbulo proximal se divide en tres segmentos; los segmentos 1 y 2 corresponden a las porciones inicial y final del tbulo contorneado proximal, el cual se contorsiona en la corteza y el segmento 3 corresponde a la pars recta, o tbulo proximal recto, que se dirige a la mdula.El tbulo proximal reabsorbe sustancias del lquido tubular y secreta sustancias al lquido tubular. As, el tbulo proximal es responsable del procesamiento inicial del filtrado glomerular. Aproximadamente dos tercios del agua y Na+ filtrados y virtualmente toda la glucosa y aminocidos filtrados se reabsorben en esta regin. Los cidos y bases orgnicos, as como drogas y metabolitos de ellas se secretan.
ASA DE HENLE
EL asa de Henle est formada por un segmento descendente y un segmento ascendente.El asa de Henle comienza en el extremo de la pars recta, donde las clulas del epitelio cbico del tbulo proximal son remplazadas por un epitelio plano, con un pequeo nmero de microvellosidades cortas y pocas mitocondrias. Este segmento del tbulo se conoce como segmento descendente delgado porque la capa de clulas epiteliales es plana, es decir, delgada, pero no significa que la luz del tbulo tenga un dimetro disminuido.En el segmento ascendente se presentan diferencias importantes entre las nefronas; en las nefronas de asa larga, el segmento ascendente comienza con un segmento de clulas de epitelio plano, llamado segmento ascendente delgado. En la unin entre la zona interna y externa de la mdula comienza el segmento ascendente grueso, este atraviesa la zona externa de la mdula (porcin medular del segmento ascendente grueso) y luego asciende a travs de la corteza (porcin cortical del segmento ascendente grueso) hasta el nivel del glomrulo de su nefrona; en las nefronas de asa corta el segmento ascendente consta por completo de clulas epiteliales cbicas, es decir, las nefronas de asa corta carecen de segmento ascendente delgado.Al igual que el tbulo proximal, el asa de Henle reabsorbe sustancias del lquido tubular y secreta sustancias a ste. En las nefronas de asa larga, hasta el 25% del agua y del Na+ filtrados se reabsorben en el asa de Henle, mientras que la urea se aade al lquido tubular. El asa de Henle tambin tiene un papel importante en el mecanismo de concentracin y dilucin de la orina.
NEFRONA DISTAL
La nefrona distal se refiere a todas las porciones del tbulo, desde el extremo del segmento ascendente grueso cortical hasta el extremo de la papila. En cada nefrona el segmento grueso ascendente vuelve a su glomrulo de origen, el tbulo contacta con su glomrulo original en el polo vascular, la regin donde entran y salen del glomrulo las arteriolas aferentes y eferentes respectivamente. En este punto de contacto se encuentra una estructura especializada, el aparato yuxtaglomerular.En la nefrona distal pueden identificarse tres segmentos principales: el tbulo contorneado distal, el tbulo conector y el tbulo colector. El tbulo colector a su vez puede subdividirse en conducto colector cortical, conducto colector medular y conducto colector papilar.El tbulo contorneado distal es un segmento relativamente corto, que se extiende hacia afuera, desde su punto de contacto con el glomrulo, hacia la superficie cortical. El tbulo conector tiene una forma algo contorneada y puede unirse con tbulos conectores adyacentes antes de su sitio de unin con el tbulo colector cortical. El tbulo colector cortical comienza como un segmento corto ms all del tbulo conector, continua como un conducto ligeramente ms grande, formado por la unin de dos segmentos o ms de este tipo, y finalmente se transforma en un conducto recto que penetra hacia la mdula. El tbulo colector medular y el tbulo colector papilar representan las extensiones de este tubo recto al interior de la mdula externa y de la papila, respectivamente. En una zona ms profunda de la papila surgen varios tbulos colectores papilares, para formar un conducto comn, llamado conducto de Bellini, que se vaca en un cliz menor.La nefrona distal es responsable de la transformacin final del lquido tubular en orina. Sus funciones incluyen la reabsorcin de Na+ y Cl- y la secrecin de H+ y K+ y tiene un papel importante en la concentracin y dilucin de la orina.
APARATO YUXTAGLOMERULAR
El aparato yuxtaglomerular (aparato YG) est formado por tres componentes: la mcula densa, las clulas mesangiales extraglomerulares y las clulas granulares. La mcula densa es una hilera de clulas epiteliales cbicas muy compactas que tapizan el tbulo en el sitio de contacto. Esta hilera de clulas define el comienzo de la nefrona distal. Las culas mesangiales extraglomerulares pueden verse como una extensin de las clulas mesangiales del glomrulo en la regin triangular, limitada por la arteriola aferente, la arteriola eferente y la mcula densa. Las clulas granulares se localizan en la regin de las clulas mesangiales extraglomerulares y en las paredes de las arteriolas aferente y eferente adyacentes. Las clulas granulares reciben su nombre debido a la existencia de grnulos de secrecin que contienen el precursor de la renina. Cuando se secreta en la luz de las arteriolas, la renina acta sobre una protena especfica del plasma, el angiotensingeno, produciendo un decapptido, la angiotensina 1. La angiotensina 1, a su vez, sufre la eliminacin de dos aminocidos por una enzima convertidora dando lugar a angiotensina 2.
Enzimaconvertidora
renina
Angiotensingeno angiotensina 1 angiotensina 2
La angiotensina 2 tiene por lo menos dos acciones importantes: estimula la secrecin de aldosterona por la corteza adrenal y es un constrictor poderoso del msculo liso arteriolar. Debido a esta ltima accin, la angiotensina 2 puede modular tanto el flujo de sangre a travs de los capilares glomerulares como la presin hidrosttica en ellos. Adems la accin vasoconstrictora de la angiotensina 2 en las arteriolas sistmicas puede aumentar la resistencia vascular perifrica y, as, mantener o elevar la presin aterial sistmica.
MTODOS EN LA FISIOLOGA RENAL
Estos mtodos pueden dividirse en cuatro categoras generales: medidas de aclaramiento, mtodos de micropuncin, perfusin de segmentos de nefrona obtenidos por microdiseccin y tejidos modelo.
MEDIDAS DE ACLARAMIENTO
El aclaramiento proporciona una manera de evaluar la eliminacin de una sustancia por los riones. Para comprender el significado de aclaramiento, considrese primero la tasa de excrecin de una sustancia X, es decir, la masa de X excretada por unidad de tiempo. Ser igual a la masa de X por unidad de volumen de orina (es decir, la concentracin de X en la orina, Ux) multiplicado por el volumen de orina excretado por unidad de tiempo (es decir, el flujo urinario, V)
Masa de X excretada = masa de X en la orina x volumen de orina = UX V Tiempo volumen urinario tiempo
Dado que una importante funcin de los riones es la extraccin de las sustancias desde el plasma, es til describir la eliminacin de X en una forma algo distinta. En lugar de hablar de tasa de excrecin de X en la orina considrese la velocidad de extraccin de X del plasma. Esto puede considerarse igual a la concentracin de X en el plasma (PX) multiplicada por el volumen de plasma, del cual X se eliminara completamente, o se depura por unidad de tiempo
Masa de x extrada del plasma = concentracin plasmtica de X x volumen de plasma depurado de X tiempo tiempo
El volumen de plasma del que X es completamente depurado por unidad de tiempo se llama aclaramiento de X y se designa como CX. Por lo tanto:
Masa de X eliminada del plasma = PX . CX tiempo
Es importante sealar que el volumen de plasma depurado de X es un volumen terico y no se puede reunir ni medir en forma directa. Esto es as porque ningn mililitro de plasma tiene todo el X eliminado por los riones, sino que cierta fraccin de X de cada mililitro de plasma se elimina. A pesar de que CX es slo un volumen terico, su valor puede calcularse a partir de volmenes medibles, dado que la conservacin de la masa implica que la velocidad de extraccin del plasma es igual a la velocidad de excrecin:
Masa de X eliminada masa de X excretada del plasma = en la orina Por lo tanto: PX CX = UX V y as: CX = UX V tiempo tiempo PX
El aclaramiento requieren slo muestras de orina y sangre perifrica, es decir, las medidas de depuracin son virtualmente no invasivas, lo que constituye una ventaja frente a los otros mtodos.
FILTRACIN Y FLUJO SANGUNEO
Aunque los riones representan slo el 0.5% del peso corporal total, reciben entre el 20% y 25% del volumen minuto del corazn. As, en un adulto promedio con un volumen minuto de 6litros/minuto el flujo sanguneo renal es mayor de 1.2 litros/minuto o 1700 litros/da. Esto significa que el volumen sanguneo total, pasa a travs del sistema vascular renal casi 300 veces por da. Mientras el flujo sanguneo renal (FSR) representa el volumen de sangre que fluye a travs del sistema vascular renal por unidad de tiempo, el flujo plasmtico renal (FPR) se refiere al flujo de plasma que pasa a travs de la vascularizacin renal. Si el hematocrito es 45%, el FPR es 55% del FSR. Por lo tanto, el FPR promedio excede 650 mil/min o 900 litros/da. En el glomrulo se filtran aproximadamente 125 ml de plasma cada minuto, es decir, el volumen de filtrado glomerular (VFG) es 125 ml/min o 180 litros/da. As, aproximadamente el 20% del FPR se filtra al espacio de Bowman. El cociente entre VFG y FPR se llama fraccin de filtracin (FF).
PROPIEDADES DE LA BARRERA DE FILTRACIN
La barrera es libremente permeable al agua y a solutos pequeos (cristaloides) como iones, glucosa y urea. Tiene permeabilidad selectiva a solutos de mayor tamao (partculas coloidales) como protenas grandes y lpidos y es casi completamente impermeable a los elementos celulares de la sangre. Entonces, excepto por la ausencia de protenas y lpidos, el filtrado glomerular es virtualmente idntico al plasma. Las sustancias con un cociente [filtrado]/[plasma] de 1 se dice que se filtran libremente. La cantidad filtrada de una sustancia de este tipo por unidad de tiempo puede calcularse con facilidad multiplicando su concentracin plasmtica por el VFG. As, para una sustancia X libremente filtrable:
Masa de X filtrada = [plasmtica X] x volumen de plasma filtrado tiempo tiempo
Px VFGEl vocablo carga filtrada se utiliza con frecuencia cuando se hace referencia a la cantidad de Px VFG. Mientras las sustancias libremente filtrables deben tener un PM 5000 daltons, la ausencia de una unin significativa con las protenas plasmticas, como la albmina, es un requisito adicional. La filtracin de ciertos iones y solutos pequeos (Ca2+, Mg2+, bilirrubina) est notablemente reducida como resultado de su unin con las protenas plasmticas. La carga filtrada de una sustancia que exhibe una unin significativa con las protenas plasmticas est dada por:
Masa de X filtrada = Px VFG . Fx donde Fx es la fraccin de sustancia que se encuentra Tiempo libre en el plasma (es decir, no unida).
La presencia de protenas en el plasma pero no en el filtrado, genera el establecimiento de un equilibrio Gibbs-Donnan entre el plasma y el filtrado. Dado que las protenas plasmticas estn cargadas negativamente, los cationes como el Na+ y el K+ tienen concentraciones algo mayores en el agua plasmtica, mientras que los aniones como el Cl- y el HCO3- tienen concentraciones ligeramente mayores en el filtrado. Sin embargo, las diferencias de concentracin atribuibles al efecto Gibbs-Donnan son pequeas y en general despreciables en la fisiologa renal.Ultrafiltrado del plasma = plasma sin protenas.
La filtracin de los solutos es inversamente proporcional a su tamao.Las molculas cargadas negativamente son menos fcilmente filtradas que aquellas cargadas positivamente.
FUERZAS IMPLICADAS EN LA FILTRACIN
El principio de Starling dice que la velocidad y direccin del movimiento de lquidos estn determinadas por el equilibrio entre la presin hidrosttica y onctica. El principio de Starling puede expresarse de la siguiente manera:
q = caudal o flujo de lquido a travs del capilar Kf = coeficiente de filtracin q = Kf [(Pc Pi) (c i)] Pc = presin hidrosttica del capilar Pi = presin hidrosttica intersticial c = presin onctica del plasma i = presin onctica intersticial
Un valor positivo de q significa filtracin neta (Pc > C), mientras que un valor negativo significa reabsorcin neta (Pc < C).En un capilar sistmico, en el extremo arterial, el equilibrio de presiones favorece a la filtracin (presin de filtracin neta = 16 mm Hg), sin embargo, en el extremo venoso tiene lugar la reabsorcin (presin de filtracin neta = -14 mm Hg), dado que la presin hidrosttica declina notablemente a lo largo de ste.
Pc (lnea que baja)c (lnea recta, constante)El resultado es que la filtracin y la reabsorcin aproximadamente se equilibran. Si se promedia la presin de filtracin neta a lo largo del capilar se obtiene una presin de filtracin neta media de 8 mm Hg. Por lo tanto, se filtra un ligero exceso de plasma al insterticio, aunque el volumen neto de filtracin en todos los capilares extrarrenales probablemente sea inferior a 2 ml/min, este volumen retorna a la circulacin sistmica por la linfa.
Las presiones que favorecen la filtracin son la Pc y i, mientras que las que favorecen la reabsorcin son la Pi y c.
Para los capilares glomerulares la ecuacin de Starling puede reescribirse como:
Kf = coeficiente de filtracin PG = presin hidrosttica del capilar glomerular VFG = Kf [(PG PB) (G B)] PB = presin hidrosttica de la cpsula de Bowman G = presin onctica del capilar glomerular B = presin onctica de la cpsula de Bowman
Dado que la cantidad de protena filtrada al espacio de Bowman es despreciable, B se aproxima a cero y la ecuacin se simplifica a:
VFG = Kf [PG PB G]
La variacin de la presin neta de filtracin en funcin de la longitud del capilar glomerular difiere, en cierta medida, de la que existe en los capilares extrarrenales. Es importante lo siguiente: 1) La presin hidrosttica de los capilares permanece relativamente constante a lo largo del capilar glomerular, en contraste con su marcada disminucin en los capilares extrarrenales. Esto puede deberse en parte a la presencia de una arteriola de elevada resistencia en el extremo eferente de los capilares glomerulares, pero no en los capilares extrarrenales.2) Dado que el lquido de filtrado en los capilares glomerulares es tan grande, la presin onctica del plasma, en la longitud de un capilar glomerular se eleva, mientras que es relativamente constante en los capilares extrarrenales. As, la filtracin tiene lugar en el extremo aferente (presin neta de filtracin = 10 mm Hg), al igual que en los capilares extrarrenales. Sin embargo en el extremo eferente el movimiento de lquido cesa (presin neta de filtracin = 0 mm Hg). Por lo tanto, se dice que el equilibrio de filtracin est presente en el extremo eferente de los capilares glomerulares. La presin neta de filtracin de los capilares glomerulares no puede determinarse con precisin, sin embargo se estima que es similar a la de los capilares extrarrenales (8 mm Hg). El hecho de que el VFG sea en promedio 125 ml/min, mientras que el flujo de filtracin neto de todos los capilares extrarrenales es menor a 2 ml/min, puede atribuirse a diferencias en el Kf. El mayor Kf de los capilares glomerulares puede deberse a las numerosas fenestras del endotelio del capilar glomerular.La presin que favorece la filtracin es la PG, mientras que las presiones que favorecen la reabsorcin son la PB y la G.
El punto a lo largo del capilar glomerular en el que se alcanza el equilibrio de filtracin esta desplazado hacia el extremo eferente si el FSR aumenta, dado que la G aumenta con menor rapidez.
PG (cte)
G
REGULACIN DEL FSR Y DEL VFG
El FSR, al igual que el flujo de sangre a cualquier otro rgano, est controlado por la diferencia de presin arteriovenosa a travs del lecho vascular (presin de perfusin) y la resistencia vascular.
FSR = P R
La presin de la arteria renal es esencialmente la misma que la de las otras arterias sistmicas (100 mm Hg), la presin hidrosttica del capilar glomerular es de aproximadamente 45 mm Hg. La gran cada de presin que ocurre en las arteriolas aferentes y eferentes identifica a estos vasos como los principales sitios de resistencia vascular renal.Factores que controlan el flujo de filtrado glomerular:Los cambios en la presin hidrosttica del capilar glomerular puede causar cambios en el VFG, la PG est controlada por las presiones y resistencias de las arteriolas aferente y eferente adyacentes. Un aumento de la resistencia de las arteriolas aferentes disminuye la PG, mientras que un aumento en la resistencia de las arteriolas eferentes aumenta la PG. Por el contrario, los cambios de resistencia de las arteriolas aferentes y eferentes en el FSR son los mismos, por ejemplo, un incremento en la resistencia arteriolar aferente o eferente disminuir el FSR. El Kf tambin afecta el VFG, por ejemplo, el Kf puede disminuir debido a enfermedades que provocan un engrosamiento de la barrera de filtracin o disminuyen su superficie al destruir capilares glomerulares; adems, muchas hormonas y otras sustancias endgenas pueden alterar el Kf posiblemente contrayendo o relajando las clulas mesangiales.Pequeos cambios en la G pueden presentarse en la deshidratacin (G aumentada) o en la hipoalbuminemia (G disminuida).El regulador ms importante es el FPR, ya que este determina la velocidad a la cual la presin onctica del plasma (G) se eleva en todo el capilar glomerular. Por ejemplo, si el equilibrio de filtracin ocurre normalmente, en un punto a dos terceras partes de la distancia entre los extremos aferente y eferente del capilar glomerular, un incremento en el FPR aumentar el VFG porque G aumenta con menos rapidez, desplazando por lo tanto ms hacia el extremo eferente del capilar el punto en el que la presin neta de filtracin se hace cero.
AUTORREGULACIN
El trmino autorregulacin, se atribuye al mantenimiento del flujo constante (FPR y VFG), a pesar de los cambios en la presin arterial sistmica, lo cual es fundamental para que se pueda llevar a cabo el mantenimiento de la cantidad y composicin normal de los lquidos corporales por parte de los riones.El FPR y el VFG permanecen casi constantes, aun en presencia de grandes cambios en la presin arterial sistmica media. En los seres humanos hay un rango de autorregulacin de aproximadamente 60 mm Hg a 130 mm Hg. A causa de la autorregulacin, la fraccin de filtracin (VFG/FPR) tambin permanece constante en este rango de presiones.Un incremento en la presin arterial debe estar acompaado de un incremento en la resistencia vascular renal para mantener un FPR constante (FSR = P/R); este incremento en la resistencia vascular renal debe ocurrir sin alterar significativamente la presin hidrosttica en los capilares glomerulares.El principal mecanismo de la autorregulacin es el mecanismo miognico, que atribuye el incremento en la resistencia vascular renal, que acompaa un incremento en la presin arterial sistmica, a la contraccin de las clulas de msculo liso de las arteriolas aferentes en respuesta al estiramiento, es decir, en respuesta al aumento de la tensin parietal, debido a que este estiramiento lleva a la apertura de canales de Ca2+ en la membrana plasmtica, dando como resultado una entrada de Ca2+ que provoca la contraccin.Un mecanismo alternativo, la hiptesis yuxtaglomerular, atribuye la autorregulacin a cambios en la tasa de secrecin de renina desde las clulas granulares, que a su vez altera la concentracin local de angiotensina 2, un potente vasoconstrictor.
REGULACIN NEURAL
Al igual que la mayora de los vasos sanguneos, los vasos del rin estn inervados exclusivamente por fibras vasoconstrictoras del simptico. En reposo, el tono simptico en los vasos renales es mnimo. La vasoconstriccin renal resultante de la estimulacin simptica provoca una reduccin, tanto en el FPR como en el VFG. Sin embargo, con una estimulacin simptica moderada el VFG no disminuye tanto como el FPR; por lo tanto, la vasoconstriccin debe comprometer tanto a las arteriolas aferentes como a las eferentes. Por lo tanto con niveles moderados de estimulacin simptica la fraccin de filtracin en realidad aumenta. Con una estimulacin simptica mayor, las disminuciones del VFG son paralelas con las del FPR; aparentemente la constriccin de las arteriolas aferentes predomina a estos niveles ms elevados de estimulacin.Es importante notar que la vasoconstriccin renal mediada por el simptico representa parte de los mecanismos del organismo para controlar la presin arterial sistmica, y no un mecanismo para regular el FPR y el VFG. Una vasoconstriccin de este tipo se inicia como respuesta refleja a una disminucin de la presin arterial sistmica, detectado a nivel de los barorreceptores del seno carotdeo y del arco artico, por ejemplo durante cambios posturales, hemorragia o sncope. Cuando disminuye la presin auricular puede iniciarse una respuesta similar, detectada por los barorreceptores localizados en las paredes de las aurculas. En todas estas situaciones la vasoconstriccin renal contribuye a incrementar la resistencia vascular perifrica total, que a su vez ayuda a restablecer la presin arterial a su valor normal. La vasoconstriccin simptica tambin puede iniciarse en respuesta a un estmulo desde el SNC, por ejemplo frente al miedo, dolor, fro, ejercicio y otras situaciones estresantes. El hecho de que el VFG no disminuye tanto como el FPR frente a niveles moderados de estimulacin simptica puede verse como una adaptacin destinada a mantener el VFG tan alto como sea posible, aunque la resistencia vascular renal deba aumentarse para controlar la presin arterial sistmica.
REGULACIN HORMONAL
La sustancia que probablemente tenga un papel fisiolgico en la regulacin del FPR y el VFG es la angiotensina 2, un potente vasoconstrictor sintetizado en respuesta a la secrecin de renina de las clulas granulares. Dado que la renina se secreta en respuesta a la disminucin de la perfusin renal y tambin en respuesta a la estimulacin simptica de las clulas granulares, al menos parte de la vasoconstriccin renal, que ocurre como resultado de una disminucin en la presin arterial sistemica, puede estar mediada por angiotensina 2.Las prostaglandinas tambin pueden tener un papel fisiolgico en la regulacin del FPR y del VFG. Los riones pueden sintetizar a varios miembros de la familia de las prostaglandinas, incluyendo la prostaglandina E2, las prostaciclinas, los leucotrienios y los tromboxanos. En trminos de los efectos sobre el FPR y el VFG, las prostaglandinas vasodilatadoras, como la prostaglandina E2, se piensa que son las ms importantes. Estas prostaglandinas pueden mediar los efectos constrictores de la estimulacin simptica o por niveles aumentados de angiotensina 2. Este papel de las prostaglandinas vasodilatadoras tambin es apoyado por el hecho de que los inhibidores de la sntesis de las prostaglandinas (por ejemplo la indometacina) pueden provocar una disminucin dramtica del FPR y del VFG en pacientes con enfermedades que disminuyen la perfusin de los riones.La bradicinina, un potente vasodilatador, puede desempear un papel similar para modular el FPR y el VFG. La hormona antidiurtica (ADH), que es de importancia capital en el mecanismo de concentracin de la orina, es un vasoconstrictor y puede disminuir el FPR y el VFG. La serotonina tambin provoca vasoconstriccin renal. La dopamina es un vasodilatador renal y puede aumentar el FPR y el VFG. El factor natriurtico auricular (FNA), que tiene un papel importante en la regulacin de la excrecin de Na+, tambin es un vasodilatador y puede aumentar el VFG. El embarazo puede aumentar el FPR y el VFG hasta en un 50%. La ingestin de una alimentacin rica en protenas puede aumentar el FPR y el VFG hasta en 30%, efecto que puede estar mediado por las prostaglandinas y ser imitado al infundir aminocidos por va intravenosa.
DIFERENCIAS INTRARRENALES EN EL FSR Y EN EL VFG
Aproximadamente el 90% del FSR total perfunde a la corteza; solo 10% perfunde a la mdula, el flujo sanguneo reducido en la mdula aparentemente se debe a la resistencia vascular relativamente elevada de los vasa recta.La existencia de un flujo sanguneo medular bajo tiene importancia considerable en el mecanismo concentrador de la orina y probablemente explica, al menos en parte, la susceptibilidad aumentada de la mdula renal a la lesin hipxica en las enfermedades.
CONSUMO RENAL DE OXGENO
Los riones difieren de la mayora de los otros rganos, ya que en ellos el flujo sanguneo no est relacionado con las demandas de oxgeno.Dado que el flujo de los riones es tan alto en relacin con sus necesidades de oxgeno, la diferencia arteriovenosa renal de oxgeno es bastante baja, aproximadamente 1,7 ml O2/100 ml de sangre, en comparacin con 4 O2/100 ml 5 O2/100 ml de sangre para el organismo en su conjunto. La relacin entre el FSR, la diferencia arteriovenosa de oxgeno y el consumo renal de este elemento es expresada por la ecuacin de Fick:
FSR = QO2 = consumo de oxgeno en el rion (RAO2 RVO2) diferencia arteriovenosa renal de oxgeno
Los riones son nicos con referencia a que los cambios en el flujo de sangre estn acompaados por cambios paralelos en el consumo de oxgeno y la diferencia arteriovenosa de oxgeno sigue siendo la misma. La explicacin para este comportamiento poco habitual es que un cambio en el FSR en general est acompaado por un cambio paralelo en el VFG, y as en la cantidad de iones y otros solutos que deben reabsorberse. A su vez, la absorcin de solutos requiere oxgeno para generar energa, entre el 75% y el 85% del consumo de oxgeno renal se utiliza para mantener la reabsorcin activa de iones y otros solutos, en particular Na+.De hecho, el consumo de oxgeno renal es directamente proporcional a la cantidad de Na+ reabsorbido. Entonces, los cambios en la reabsorcin de soluto son responsables de los cambios en el consumo de oxgeno que acompaan a los cambios en el FSR.
SINTESIS DE ERITROPOYETINA
Los riones tambin cumplen la funcin de monitorear la adecuacin del envo de oxgeno a los tejidos, en general. En respuesta a la hipoxia renal, las clulas endoteliales de los capilares peritubulares sintetizan eritropoyetina, una hormona que regula la produccin de eritrocitos a partir de clulas precursoras de la mdula sea.
MEDIDA DEL VFG
El VFG puede medirse con la ayuda de ciertas sustancias que se filtran libremente en el glomrulo, pero que no son reabsorbidas o secretadas por el tbulo. La inulina puede considerarse el ejemplo prototipo, dado que esta no es extrada del filtrado por reabsorcin, ni es aadida a este por secrecin, la cantidad total filtrada se excretar en la orina. As, la tasa de excrecin ser igual a la carga filtrada:
Masa de inulina excretada = masa de inulina filtrada o UIn V = PIn VFG tiempo tiempo Si esta ltima ecuacin se sustituye en la expresin utilizada para el aclaramiento de la inulina, se obtiene el siguiente resultado:
CIn = UIn V = PIn VFG por lo tanto: CIn= VFG PIn PIn
CIn, por supuesto, puede calcularse a partir de las medidas hechas en muestras de plasma y orina. As esta ecuacin define mtodo no invasivo para medir el VFG.Como la inulina no se reabsorbe ni se secreta, el volumen de plasma depurado de inulina por unidad de tiempo (CIn) es igual al volumen filtrado por unidad de tiempo. Adems, no es txica, no se une a las protenas plasmticas, no se metaboliza ni se sintetiza en los riones y no tiene efectos sobre el VFG. Aunque la inulina se considera la sustancia prototipo para la medida del VFG, se han identificado otras sustancias adecuadas, principalmente el manitol y el iotalamato. La prctica ms comn para estimar el VFG en la clnica es utilizar la sustancia endgena creatinina; esta es filtrada en el glomrulo y secretada a la luz del tbulo por las clulas epiteliales del tbulo proximal. Sin embargo, a niveles plasmticos normales en los seres humanos, la cantidad de creatinina secretada es slo aproximadamente 10% a 15% de la cantidad filtrada. Esto significa que:
Masa de creatinina masa de creatinina excretada = filtrada o UCr V PCr VFG tiempo tiempo
Como aproximacin, entonces, la creatinina se comporta como las sustancias que solamente se filtran, como la inulina. Por lo tanto, el aclaramiento de la creatinina se puede utilizar en la clnica para obtener un valor aproximado del VFG:
CCr = UCr V PCr VFG VFG PCr PCr
Mientras el CCr representa una aproximacin clnica til para el VFG, puede obtenerse informacin importante sobre el VFG de una medida simple de PCr. Esto ocurre porque la velocidad de excrecin de creatinina debe igualar a la velocidad de su produccin y, por lo tanto, tiende a ser constante de un da a otro en estado estacionario, es decir: UCr V = constante. Dado que UCr V = PCr CCr, la cantidad de PCr CCr tambin tiende a ser constante: PCr CCr o PCr VFG = constante.Por lo tanto, la PCr es inversamente proporcional al VFG.La PCr cambia poco cuando el VFG cae de su valor normal, de 125 ml/min, a un valor tan bajo como 60 ml/min; sin embargo, la PCr comienza a aumentar notablemente con disminuciones ulteriores en el VFG. Esto significa que la PCr es un indicador sensible del VFG slo a VFG bajos. Por lo tanto la PCr no es til para detectar disminuciones pequeas o aun moderadas en el VFG, debido a una disminucin pequea en la funcin renal, pero es extremadamente til para detectar las grandes disminuciones en el VFG que se ven en la disfuncin renal grave.
MEDIDA DEL FPR
La ecuacin de Fick proporciona la base para la medida del FPR. Para esta aplicacin, la ecuacin de Fick se escribe como:
FPR= QX Donde QX es la velocidad a la cual la sustancia X es consumida por los (PRAX PRVX) riones y PARAX y PRVX, representan las concentraciones de X en el Plasma renal arterial y venoso, respectivamente. Para las sustancias que no se metabolizan y no son sintetizadas por los riones, la velocidad a la que la sustancia es consumida por estos puede igualarse a la velocidad de excrecin, UXV. Ms aun, si la sustancia no es consumida por otros tejidos distintos de los riones, la PRAX puede igualarse a PX, as:
FPR = UXV (PX PRVX)
Los niveles plasmticos en las venas renales de PRVX no son fcilmente obtenibles en los estudios clnicos rutinarios. La sustancia ideal para una determinacin del FPR no invasiva con la ecuacin de Fick, sera entonces aquella que tuviese una concentracin en el plasma venoso renal de cero, es decir una sustancia que se elimine totalmente del plasma arterial renal por accin de los riones. Para una sustancia de este tipo, la ecuacin de Fick se reduce a la ecuacin de aclaramiento:
FPR = UXV Qu sustancia tendra una concentracin (PX 0) = CX de cero en el plasma venoso renal?
Dado que solo se filtra 20% del FPR, la sustancia no se depurara completamente del plasma renal slo por filtracin; tambin sera necesario aadirla al lquido tubular por secrecin. Ms aun, esta secrecin debera ser lo suficientemente eficiente como para eliminar virtualmente toda la sustancia que no se filtr. La sustancia ms prxima a satisfacer estos criterios es el p-aminohipurato (PAH). El PAH se filtra en el glomrulo y es secretado por las clulas del epitelio del tbulo proximal, es decir, es transportado desde los capilares peritubulares que rodean al tbulo proximal hacia la luz del tbulo. Ms aun, a bajos niveles plasmticos de PAH, es tan eficiente que virtualmente todo el PAH de los capilares peritubulares se secreta a la luz del tbulo proximal. Pero aun as, el PAH no se depura completamente del plasma renal, porque entre el 10% y el 15% FPR total irriga porciones de los riones que no pueden eliminar el PAH (o cualquier otra sustancia), por ejemplo la cpsula renal, la pelvis renal, la grasa perirrenal, la mdula y la papila. As, la concentracin de PAH, o de cualquier otra sustancia, en el plasma venoso renal no es cero. Se utiliza la llamada fraccin de extraccin (EPAH) para representar a la fraccin de PAH que es depurada o extrada del plasma renal en un nico paso a travs de los riones:
EPAH = (PPAH PRVPAH) Dado que entre el 10% y el 15% del FPR total no es depurado de PPAH PAH, el EPAH tiene un valor de 0.85 a 0.90.
Mientras el CPAH slo se aproxima al FPR total, mide con precisin el flujo plasmtico a las regiones de los riones que pueden eliminar PAH, el llamado flujo plasmtico renal efectivo (FPRE):
CPAH = UPAHV = FPRE PPAHAl combinar las ecuaciones de FPR y EPAH, es evidente que: FPRE = FPR . EPAHFSR = FPR Donde Hct es el hematocrito. (1 Hct)
CLCULO DE LA FRACCIN DE FILTRACIN
Una vez que el FPR y el VFG se han determinado, puede calcularse rpidamente la fraccin de filtracin. El valor ms preciso de la fraccin de filtracin se obtendra de la siguiente manera:
FF = CIn Donde FPR se determina con la ecuacin de Fick. Sin embargo, para propsitos FPR clnicos, una aproximacin til es: FF = CCr CPAH
Determinacinms precisaAproximacin til confines clnicos
VFGCInCCr
FPREcuacin de FickCPAH
INTRODUCCIN A LA FUNCIN TUBULAR
La porcin tubular de la nefrona modifica el contenido del filtrado glomerular, de modo tal que la orina final contiene solamente aquellos constituyentes que deben excretarse para preservar el volumen y composiciones normales de los lquidos corporales. Esta modificacin implica tanto la reabsorcin tubular, el proceso por el que el agua y otras sustancias esenciales del filtrado glomerular se recuperan y vuelven al organismo por medio de los capilares peritubulares, como la secrecin tubular, proceso por el cual las sustancias de los capilares peritubulares se transportan a travs del epitelio tubular y se aaden al lquido tubular.
REABSORCIN TUBULAR
Para una sustancia X, la cantidad reabsorbida por unidad de tiempo puede calcularse como la diferencia entre la carga filtrada y la excretada:
Masa de X reabsorbida = masa de X filtrada - masa de X excretada = TX = PX VFG - UXV tiempo tiempo tiempo
Donde TX representa la velocidad de reabsorcin tubular de X. Otro concepto importante es el de reabsorcin fraccionada, es decir, la fraccin de carga filtrada que se reabsorbe, la cual se puede calcular como:
Reabsorcin fraccionada = TX La reabsorcin de ciertos constituyentes esenciales, como PX VFG la glucosa y los aminocidos, es virtualmente completa, es decir, toda la carga filtrada se reabsorbe.
Para una sustancia filtrada y reabsorbida: CX < VFG, o dado que el VFG puede ser igual al aclaramiento de la inulina: CX < CIn. Este resultado es el esperado dado que el volumen del plasma depurado de una sustancia que se filtra y luego reabsorbe es menor que el que ser depurado de una sustancia que solo se filtra, como la inulina. Los mecanismos de transporte implicados en la reabsorcin tubular de sustancias pueden clasificarse en general como activos o pasivos. En la reabsorcin pasiva una sustancia difunde de la luz del tbulo al capilar peritubular a favor de un gradiente osmtico, elctrico y/o de concentracin. No se requiere energa metablica en forma directa, aunque puede haberse necesitado para establecer el gradiente inicial.Los transportes activos pueden ser primarios o secundarios; la reabsorcin activa primaria ocurre contra un gradiente elctrico y/o de concentracin y requiere el gasto de energa metablica. El transporte activo secundario es un tipo particular de difusin facilitada que utiliza un transportador capaz de movilizar dos o ms sustancias en el mismo sentido (cotransportador o simport) o en sentido contrario (contransportador o antiport), utilizando el gradiente de difusin de uno de ellos. Este gradiente es generado y mantenido por una bomba (transporte activo primario).
REABSORCIN ACTIVA: REABSORCIN LIMITADA POR Tm
Para muchas sustancias reabsorbidas en forma activa, la velocidad de reabsorcin tiene un lmite superior finito. La velocidad mxima que puede lograrse se conoce como transporte mximo (Tm). La glucosa, los aminocidos, el fosfato y el sulfato son ejemplos de sustancias que exhiben una reabsorcin limitada por Tm. Ms aun, la afinidad del sistema de transporte por muchas sustancias con reabsorcin limitada por Tm es tan alta que toda la carga filtrada se reabsorbe desde el lquido tubular, en tanto el sistema de transporte no est saturado. Por ejemplo, la carga filtrada de glucosa es virtualmente completa si la carga filtrada no satura el sistema de transporte , es decir, la velocidad de reabsorcin iguala a la carga filtrada:
Masa de glucosa reabsorbida = masa de glucosa filtrada o TG = PGVFG tiempo tiempo
En estas condiciones resulta evidente que no se excreta glucosa. Sin embargo, cuando el sistema de transporte est saturado, el exceso de carga se excretara. Bajo tales condiciones el sistema de transporte estar operando a su velocidad mxima, es decir:
Masa de glucosa reabsorbida = TmG La velocidad de excrecin por lo tanto, ser: tiempo UGV = PGVFG - TmG
Entonces es posible definir la concentracin plasmtica en la que comienza la glucosuria. Esta concentracin plasmtica crtica se llama umbral renal para la glucosa (PThG). Tericamente, el PThG representa aquella concentracin de plasma a la cual la carga filtrada satura exactamente el sistema de transporte, es decir: PThG VFG = TmG o PThG = TmG VFG
CURVAS DE TITULACIN DE GLUCOSA
Las caractersticas importantes de las curvas de titulacin de glucosa son las siguientes:1) Carga filtrada en funcin de PG. Dado que el VFG es constante, la grfica de carga filtrada (PGVFG) en funcin de PG es una lnea recta, cuya pendiente es el VFG. (AZUL)2) Velocidad de reabsorcin tubular en funcin de PG. A una PG baja, toda la carga filtrada se reabsorbe. A una PG ms elevada, el sistema de transporte se satura y reabsorbe glucosa a su velocidad mxima, TmG. (ROJA)3) Velocidad de excrecin en funcin de PG. Dado que toda la glucosa se reabsorbe a una PG baja, la glucosa estar virtualmente ausente en la orina. Sin embargo, una vez que el sistema de transporte se satura, el exceso de carga filtrada se excreta. En este ejemplo, en el que el VFG es de 125 ml/min, la glucosuria aparece primero a una PG de aproximadamente 16 mmol/litro (300 mg/dl), es decir, el umbral renal para la glucosa es 16 mmol/litro. Cuando PG excede el umbral renal, la curva de velocidad de excrecin en funcin de PG corre paralela con la curva de carga filtrada en funcin de PG, es decir, es una lnea recta cuya pendiente es el VFG. (VERDE)
Estas curvas de titulacin son ideales, ya que muestran un cambio abrupto cuando la velocidad de reabsorcin alcanza su nivel mximo. Las curvas de titulacin de glucosa reales muestran un cambio ms gradual en esta regin. Esta caracterstica de las curvas de titulacin renal se llama explayamiento (splay), en general se explica por una capacidad de reabsorcin de glucosa diferente segn las nefronas involucradas, porque algunos se saturan antes que otros. A causa de este explayamiento, el umbral renal para la glucosa es aproximadamente 11 mmol/litro (200 mg/dl).
Tambin es de inters examinar el efecto de la PG en el aclaramiento de la glucosa. A una PG baja, toda la carga filtrada se reabsorbe, de modo que no hay plasma depurado de glucosa:
CG = UGV = 0 PG
Una vez que el umbral renal para la glucosa es excedido y comienza a parecer la glucosuria, el aclaramiento de glucosa aumenta gradualmente. CG se aproxima asintticamente a CIn a una PG alta, pero CG siempre es menor que CIn, dado que cierta cantidad de glucosa siempre se reabsorbe.
REABSORCIN PASIVA: UREA
En muchas partes de la nefrona, la urea se reabsorber pasivamente siempre que su concentracin en el lquido tubular exceda la del lquido peritubular que lo rodea. Dado que la reabsorcin de urea ocurre como consecuencia de la reabsorcin de agua, la reabsorcin de urea vara notablemente con el flujo de orina (es decir, con el volumen de agua que no se reabsorbe). Esta dependencia de la reabsorcin de V es una caracterstica nica de las sustancias reabsorbidas pasivamente, al igual que la urea. La cantidad excretada de urea se incrementa al aumentar V (debido a la reabsorcin disminuida), de modo tal que el aclaramiento de la urea aumenta a medida que aumenta la V.
SECRECIN TUBULAR
La secrecin tubular es el proceso por el cual las sustancias de los capilares peritubulares se transportan a travs del epitelio del tbulo a la luz de este. As, mientras en la reabsorcin tubular las sustancias se recuperan del lquido tubular, en la secrecin tubular se aaden sustancias al lquido tubular. Es importante distinguir entre secrecin y excrecin, el proceso completo por el cual los riones componen la orina final. Por ejemplo, podra filtrarse una sustancia a travs del glomrulo, reabsorberse parcialmente en una regin de la nefrona o ms, no secretarse nunca y aun excretarse.Para una sustancia X la cantidad secretada por unidad de tiempo puede calcularse como la diferencia entre la velocidad de excrecin y la carga filtrada:
Masa de X secretada = masa de X excretada masa de X filtrada TX = UXV - PXVFG tiempo tiempo tiempo
Para una sustancia que se filtra y se secreta: CX > VFG CX > CInEste resultado es el esperado, dado que un mayor volumen de plasma ser depurado de una sustancia que se filtra y luego se secreta que de una sustancia, como la inulina, que solo se filtra.Al igual que los mecanismos para la reabsorcin tubular, los mecanismos implicados en la secrecin tubular pueden ser clasificados en forma general como activos o pasivos.
SECRECIN ACTIVA, LIMITADA POR Tm
Ciertas sustancias que se secretan activamente exhiben una secrecin Tm limitada, es decir, la velocidad de secrecin tiene un lmite superior finito llamado transporte mximo (Tm). El p-aminohipurico (PAH), el yodohipurico y la penicilina se encuentras entre las sustancias cuya secrecin es limitada por Tm. Algunas de estas sustancias tambin muestran un fenmeno que guarda cierta analoga con el fenmeno de umbral para la reabsorcin. Especficamente, la afinidad del sistema de transporte para algunas sustancias secretadas es tan alta, que esencialmente, toda la sustancia entregada a los capilares peritubulares es secretada en el lquido tubular, en tanto el sistema de transporte no est saturado. Por ejemplo, el PAH y el yodohipurato se eliminan casi completamente de los capilares peritubulares cuando el sistema de transporte no est saturado.La velocidad de envo del PAH a los capilares peritubulares est dada por:
Masa de PAH entregada a los volumen de plasma entregado a los capilares peritubulares = PPAH x capilares peritubulares tiempo tiempo
El flujo de plasma entregado a los capilares peritubulares puede calcularse por aproximacin, restando el VFG del FPR total, es decir: Esta ecuacin es una aproximacin, Volumen de plasma entregado a los porque como sabemos, aproximadamente capilares peritubulares FPR VFG 10% al 15% del FPR total irriga regiones de tiempo los riones en las que no hay filtracin o secrecin de PAH.As, el flujo de plasma entregado a os capilares peritubulares est dado con mayor precisin por:
Volumen de plasma entregado a los Donde FPRE, es el flujo plasmrico renal capilares peritubulares = FPRE VFG efectivo, es decir, el flujo plasmtico tiempo renal a las regiones de los riones que pueden eliminar PAH.Por lo tanto, la primer ecuacin puede expresarse como:
Masa de PAH entregada a los capilares peritubulares = PPAH (FPRE VFG) tiempo
Como se expres previamente, cuando el mecanismo secretor no est saturado, esencialmente todo el PAH entregado a los capilares peritubulares se secreta:
Masa de PAH entregado a los Masa de PAH secretado = capilares peritubulares TPAH = PPAH (FPRE VFG) tiempo tiempo
En tales condiciones, entonces, el PAH se elimina casi completamente del plasma que pasa por los riones. Aproximadamente el 20% del PAH entregado a los riones entra al lquido tubular por la filtracin glomerular, el 80% restante entra a los capilares peritubulares y se secreta, una vez que el mecanismo secretor se satura, la velocidad de secrecin de PAH se fija en su valor mximo, TmPAH.
CURVAS DE TITULACIN DE PAH
Las caractersticas importantes de estas curvas son las siguientes: 1) Carga filtrada en funcin de PPAH. Dado que el VFG es constante, la curva de carga filtrada (PPAHVFG) en funcin de PPAH es una lnea recta, cuya pendiente es el VFG.2) Velocidad de secrecin tubular en funcin de PPAH. A PAH bajo, donde el sistema de transporte es capaz de secretar esencialmente todo el PAH entregado a los capilares peritubulares, la velocidad de secrecin es considerablemente mayor que la carga filtrada. A PPAH ms alta, el sistema de transporte se satura y secreta PAH a su velocidad mxima, TmPAH. En este ejemplo, TmPAH tiene un valor de 0.4 mmol/min (80 mg/min) que es el valor promedio normal en los hombres.3) Velocidad de excrecin en funcin de PPAH. La curva se eleva en forma pronunciada a una PPAH baja. Una vez que el sistema de transporte se satura, sin embargo, la curva de velocidad de excrecin en funcin de PPAH es paralela con la carga filtrada en funcin de PPAH, es decir, es una lnea recta cuya pendiente es el VFG.Debemos notar que las curvas de titulacin de PAH muestran un cambio gradual, as que tambin presentan el fenmeno de explayamiento.
A PPAH baja esencialmente todo el PAH es eliminado del FPRE por los riones, todo el FPRE es depurado de PAH, es decir: CPAH = FPRE.A PPAH alta, cuando el sistema de transporte est saturado parte del PAH de los capilares peritubulares no se secretar, es decir, hay un volumen plasmtico menor depurado de PAH.El CPAH se aproxima asintticamente a CIn a alta PPAH, pero el CPAH siempre es mayor que el CIn, dado que siempre se secreta parte del PAH.
TRANSPORTE BIDIRECCIONAL
Ciertas sustancias se transportan a travs del epitelio tubular en ambas direcciones, es decir, se filtran y luego se reabsorben y se secretan. Para propsitos cuantitativos, una sustancia que sufre este transporte bidireccional se categoriza de acuerdo con la exigencia de una reabsorcin neta o de una secrecin neta.El manejo renal de K+ representa un ejemplo importante de transporte bidireccional. Mientras el K+ sufre una reabsorcin neta, puede haber secrecin neta bajo ciertas condiciones, tales como la ingesta de una dieta elevada en K+. Otra sustancia endgena que exhibe transporte bidireccional es el cido rico. A diferencia de lo que ocurre con el K+, el cido rico siempre sufre reabsorcin neta.
CIDOS Y BASES ORGNICOS DBILES
El epitelio del tbulo proximal contiene un sistema de transporte activo especial, capaz de secretar diversos cidos orgnicos y otro, capaz de secretar diversas bases orgnicas. El PAH es un ejemplo de sustancia secretada por el sistema de transportes para cidos orgnicos. Es importante enfatizar que estos sistemas de transporte slo secretan molculas orgnicas que estn ionizadas en el plasma. Mientras que los cidos orgnicos fuertes y las bases estn completamente ionizadas en el plasma (pH = 7.4), esto no es necesariamente cierto para los cidos y bases orgnicos dbiles, que pueden existir en parte o an principalmente en forma no ionizada a pH 7.4. Muchos cidos y bases orgnicos dbiles son lo suficientemente liposolubles como para difundir pasivamente a travs del epitelio del tbulo.Al igual que otros constituyentes plasmticos, una base o cido dbil, que se filtra libremente, estarn presentes en el filtrado con una concentracin virtualmente idntica a la del plasma. A medida que el agua se reabsorbe, su concentracin en el lquido tubular exceder a la del plasma, es decir, se genera un gradiente de concentracin que favorece la reabsorcin. Sin embargo, dado que slo la forma no ionizada es lo suficientemente liposoluble como para difundir a travs del epitelio tubular, es el gradiente de concentracin de la forma difusible no ionizada (no el gradiente de concentracin total) lo que tiene importancia.En cualquier regin dada de la nefrona, el gradiente de concentracin de la forma no ionizada puede favorecer la reabsorcin o bien la secrecin, lo que depender de la relacin entre el pH del lquido del tbulo en esa regin y el pka del cido o la base. Los cidos y bases orgnicos dbiles, por lo tanto, pueden mostrar transporte bidireccional.Un lquido tubular cido favorece la reabsorcin de los cidos orgnicos dbiles, mientras que un lquido tubular alcalino favorece su secrecin y por lo tanto la excrecin final. El fenmeno por el cual la forma no ionizada de una molcula puede difundir fcilmente a travs del epitelio del tbulo, mientras la forma ionizada no puede hacerlo, se llama difusin no inica o atrapamiento de la difusin.Un lquido tubular alcalino favorece la reabsorcin de las bases orgnicas dbiles, mientras un lquido tubular cido favorece su secrecin y por lo tanto la excrecin final. Muchas drogas y metabolitos de las drogas son cidos y bases orgnicos dbiles. La orina normalmente es cido (pH promedio < 6.0), lo que favorecera la reabsorcin de drogas cidas y la excrecin de drogas bsicas.
EXCRECIN FRACCIONADA YREABSORCIN FRACCIONADA
La excrecin fraccionada de una sustancia X (EFX) puede definirse como la fraccin de la cantidad total de X en el filtrado glomerular que aparece en la orina final. As:
Masa de X excretada Dividiendo el numerador y el denominadorEFX = masa de x filtrada por el tiempo se obtiene: UXV EFX = PXVFG
La fraccin de la cantidad total de X filtrado que permanece en el lquido del tbulo, en cierto punto de la nefrona, se llama entrega fraccionada de X (FDX), dado que representa la fraccin de X en el filtrado que es entregada en ese punto.
LTX/PX Donde LT se refiere a la concentracin en el lquido tubular en el punto en FDX = LTIn/PIn cuestin.
Una vez que se conoce la excrecin fraccionada de una sustancia X, puede calcularse rpidamente la reabsorcin fraccionada, dado que:
UX/PXReabsorcin fraccionada = 1 EFX = 1 UIn/PIn
En forma similar, una vez que se conoce la entrega fraccionada de una sustancia X en un punto en particular de la nefrona, la fraccin reabsorbida hasta ese punto puede calcularse fcilmente:
LTX/PXFraccin reabsorbida hasta el punto de la muestra = 1 LTIn/PIn
EXCRECIN FRACCIONADA Y REABSORCIN FRACCIONADA DE AGUA
La excrecin fraccionada de agua (EFH2O) se define por la expresin:
Volumen de agua excretado Dividiendo el numerador y el denominador por el EFH2O = volumen de agua filtrado tiempo se obtiene: V EFH2O = IFG
Por un razonamiento anlogo, puede calcularse que la fraccin del volumen total de agua filtrada, que permanece en el tbulo en cierto punto de la nefrona, en la entrega fraccionada de agua (FDH2O) est definida por:
1 La reabsorcin fraccionada de agua puede calcularse a partir de la excrecinFDH2O = LTIn/PIn fraccionada de la siguiente manera: 1 Reabsorcin fraccionada = 1 EFH20 UIn/PIn
En forma similar, la fraccin de agua filtrada reabsorbida hasta cierto punto de la nefrona puede calcularse de la entrega fraccionada 1Fraccin reabsorbida hasta el punto de la muestra = 1 FDH2O 1 LTIn/PIn
EL TBULO PROXIMAL
Entre sus funciones importantes se encuentra la reabsorcin de aprox. de Na+ y agua filtrados, la reabsorcin de virtualmente toda la glucosa y AA filtrados y la secrecin de cidos y bases orgnicas.PERMEABILIDAD AL AGUA
Un determinante importante de la funcin del tbulo proximal es la alta permeabilidad al agua, que evita el establecimiento de un gradiente osmtico apreciable a travs del epitelio del tbulo proximal. Fisiolgicamente, esto significa que la reabsorcin de cualquier soluto por el tbulo proximal dar como resultado la reabsorcin de agua, de modo que la osmolaridad del lquido tubular sigue siendo aproximadamente igual a la del lquido peritubular. En realidad, la osmolaridad del liquido peritubular es entre 1 mosmol/kg de H2O y 5 mosmol/Kg de H2O superior a la del liquido tubular, debido a la acumulacin del soluto reabsorbido en el espacio peritubular. El liquido peritubular que rodea al tbulo proximal, es decir, el liquido peritubular cortical tiene una osmolaridad igual a la del plasma; as, el liquido del tbulo proximal es ligeramente hipotnico.
REABSORCION DE Na+
Aproximadamente 2/3 partes del Na+ filtrado se reabsorben en el tbulo proximal. El Na+ debe estar acompaado por un anion para mantener la electroneutralidad; aprox. 75 % est acompaado por Cl- , mientras el 25% restante est acompaado por HCO3- . Aunque la reabsorcin de cualquier soluto por el tubulo proximal dara como resultado la reabsorcin de agua, el Na+ y los aniones acompaantes son los principales solutos reabsorbidos por el tubulo proximal y, por lo tanto, son los principales responsables de generar la fuerza osmtica impulsora para la reabsorcin de agua; osea la reabsorcin de las 2/3 partes del Na+ filtrado es la responsable de los 2/3 de agua filtrada. Se han identificado 3 mecanismos principales de transporte: el cotransporte Na+/ soltuto, el intercambio Na+/H+ (antiporte) y el transporte de Na+ impulsado por Cl-.En primer lugar se deben considerar ciertas caractersticas bsicas que comparten el cotransporte Na+/soluto y el mecanismo de intercambio Na+/H+. Ambos son ejemplos de reabsorcin activa de Na+. En estos mecanismos el Na+ entra a las clulas del epitelio del tubulo a travs de su superficie apical y luego es expulsado activamente a travs de la superficie basolateral de la celula. La entrada de Na+ esta mediada por transportados (aunque los transportadores implicados son diferentes en cada mecanismo) y es impulsada por un gradiente electroqumico. La salida activa de Na+ es llevada a cabo por la bomba Na+/K+ ATPasa, que tambin genera el gradiente electroqumico para la entrada de Na+. Asi la energa para el cotransporte y para el antiporte, incluyendo al gradiente electroqumico para la entrada de Na+, proviene del ATP por medio de la bomba Na+/K+ ATPasa.
COTRANSPORTE Na+/SOLUTO
La entrada apical en el mecanismo de cotransporte Na+/soluto esta acoplada al transporte de una de varias molculas importantes