Embed Size (px)
Citation preview

41. 聴覚を獲得するメカニズムの研究
小田 洋一
Key words:内耳,有毛細胞,耳石,マウスナー細胞,ゼブラフィッシュ
名古屋大学 大学院理学研究科脳機能構築学グループ
緒 言
動物の聴覚は他者とのコミュニケーションや外界の状況を認識するための重要な情報手段である.我々脊椎動物が音を聞くためには,内耳の有毛細胞で音の微小な振動を電気信号に変換する過程と,脳で音信号の特徴を抽出する過程が必要である.聴覚に関してはこれまで主に哺乳類,鳥類,両生類を用いて膨大な研究がなされていて,内耳有毛細胞の機械刺激受容チャネル候補分子,有毛細胞から蝸牛神経核への聴覚回路形成,蝸牛神経核細胞の発達などが明らかにされた.ところが,有毛細胞の先端に存在するとされる機械受容チャネルは,いまだに未同定でありそのメカニズムも解明されていない.また,胚の中枢神経細胞への電気生理学的アプローチはこれまで困難を極め,発達初期の聴覚回路の構築や機能の発達について本質的な問題が未解明のまま残されていた.我々はモデル脊椎動物をして近年注目されているゼブラフィッシュを用いて,聴覚の発達過程を生理学的・形態学的に解析し,聴覚獲得のキィとなる候補因子を明らかにするに至り,本研究でその実体を突き止めようと計画した. ゼブラフィッシュは音に敏感で, 高等脊椎動物と同じように内耳の有毛細胞が音を電気信号に変換する. 音の情報は聴神経から後脳に左右一対存在するマウスナー(M)細胞に直接入力する. 我々は運動中のゼブラフィッシュからニューロン活動をカルシウムイメージングによって記録し, M 細胞が強力な聴覚入力を受けて最も早い聴覚性逃避運動を駆動することを明らかにした 1). さらにゼブラフィッシュ胚の M 細胞のホールセル記録および内耳有毛細胞から音応答電位を記録する手法を世界に先駆けて確立した 2).本研究では,発達過程でどのようにして聴覚を獲得するかを理解するために,ゼブラフィッシュの胚や仔魚を対象にして,内耳での「音の受容」と聴覚中枢ニューロンでの「音の特徴抽出」を達成するメカニズムの機能的発達を解析した.
方法、結果および考察
1.聴覚誘導性逃避運動の獲得 内耳で音が電気信号に変換されても,その情報が正しく脳に伝わり処理されなければ認知されない.音情報が動物に正しく認知されたかを調べるには,音に対する動物の応答を見るのが良い.ゼブラフィッシュの聴覚に依存した応答の中で逃避運動はその基本神経回路(図1A)が感覚入力から運動出力まで良く理解されていて,聴覚の発達を調べるにはもっともすぐれた標本のひとつである. 我々はこれまでに,ゼブラフィッシュが聴覚刺激からすばやく逃げる逃避運動が,後脳に左右一対存在する巨大な網様体脊髄路ニューロンである M 細胞によって駆動されることを,逃避運動中の M 細胞の活動をカルシウムイメージングによって明らかにした 1).いつから音に対する応答は現れるのであろうか? 受精後2~7日のゼブラフィッシュ仔魚に音刺激を与えて調べると,逃避運動が観察されるのは受精後 75 時間以降であった(図1B).それ以前は触刺激に対しては明確な逃避運動を示すが音刺激には全く応答しない.この発達初期に M 細胞と逃避運動を同時計測すると触刺激に対して M 細胞が発火していた.M 細胞を破壊すると触刺激に対するもっとも早い逃避運動が失われた.その後,聴覚性の逃避運動が現れる頃になると M 細胞は音刺激に対して発火するようになる一方,これまで応答していた触刺激に対しては発火しなくなる.すなわち,M 細胞は発達初期には触刺激に対して発火し,受精後 75 時間以降では主要入力を聴覚に切り替えて発火してより早い逃避運動を駆動すると結論した 3).
1
上原記念生命科学財団研究報告集, 27 (2013)

図 1. ゼブラフィッシュの逃避運動回路と音刺激に対する逃避運動の発達.A.ゼブラフィッシュの逃避運動回路.ゼブラフィッシュは音に敏感で,大きな音に対して素早く体を曲げて逃げる.音は内耳の有毛細胞でとらえられ,聴神経を介して後脳に左右一対存在するマウスナー細胞に伝えられる.マウスナー細胞が活動すると反対側の脊髄の運動細胞や介在細胞が活動し,短潜時で胴筋が収縮し反対側へ体を屈曲させ,逃避運動が開始する.B.音刺激に対する逃避運動の発達.受精後 75 時間以前のゼブラフィッシュは触刺激には逃避運動を示すが,音刺激には全く応答しない.75 時間から 100 時間の間に音刺激に対して逃避する確率が急速に上昇する.
2.聴覚回路の発達 M 細胞への聴覚入力は発達初期にどのように変化するのであろうか? ゼブラフィッシュ生体内の M 細胞をホールセル記録して,音刺激に対する応答を調べると(図2A),受精後 90 時間(3日齢)では,音刺激から2ミリ秒の潜時
(刺激から応答の開始までの時間)で鋭い内向きのシナプス電流が得られた.これは内耳の有毛細胞から聴神経(第VIII 神経)を経て M 細胞に至る経路を経て誘発されたものである.一方,受精後 40 時間までは強い音刺激を与えても,M 細胞には応答は得られない.最初に応答が観察されるのは受精後 46 時間である.しかし,応答の潜時は長く
(約 10 ミリ秒)振幅も小さくて波形は安定しない.その後,潜時は急激に短くなり振幅も増大してくる 2). M 細胞が音に初めて応じる時期(受精後 46 時間)は何で決まるのだろうか? そもそも内耳の有毛細胞から聴神経を経て M 細胞に至る回路はいつできるのであろうか? 我々はまず内耳の有毛細胞の出現と発達を形態学的・生理学的に解析し,最初の有毛細胞が受精後 22 時間に生まれることを見出した 4).サカナの内耳有毛細胞は音の振動を受けると,ステレオシリアと呼ばれる繊毛の先端に存在する耳石との間にずれが生じ,ステレオシリアの頂上に発現するとされる機械受容チャネルが開いて陽イオンが流れ込み脱分極をする.大変興味深いことに,最初のステレオシリアは耳石を先端につなぐ Tether cells に現れた.ステレオシリアが形成された有毛細胞は1時間後には繊毛の機械刺激に対して応答した.これまでに,M 細胞が受精後 7.5 時間で前駆細胞から分化してくることや聴神経を含む第 VIII 神経が受精後 23 時間には M 細胞とシナプス結合していることは知られていた.聴神経と内耳有毛細胞との結合について,第VIII 神経に緑色蛍光タンパク(GFP)を発現するトランスジェニックゼブラフィッシュを利用して調べると,受精後27 時間ですでに第 VIII 神経は有毛細胞内耳有毛細胞にシナプス構造を作って結合していた 2).この時期はまだ音には応じないが,有毛細胞に水流刺激を当てたり,微小なガラス棒で有毛細胞の繊毛を長い毛の方向へ倒してやると,M細胞でシナプス電位が記録された.すなわち,音刺激よりずっと大きな機械刺激なら,それを受容し中枢に伝えうる回路がすでに受精後 27 時間にはできていることを示している.しかし,この回路は音に反応することができない.
2

図 2. マウスナー細胞 (A) と内耳有毛細胞 (B) の音刺激に対する応答の発達の発達.A. 発達過程のゼブラフィッシュの後脳マウスナー細胞から生体内ホールセル記録を行い,音刺激(500 Hz)に対する応答を調べた.最初に応答が得られたのは受精後 46 時間で,その後応答の振幅は大きくなり潜時は短くなった.B. 内耳の原器である耳胞のなかでは音刺激に対する有毛細胞の活動を反映するマイクロフォン電位が記録される.マイクロフォン電位も受精後 46 時間から観察され,その後次第に振幅が大きく潜時は短くなった.これらの応答は有毛細胞の繊毛の先端に存在する機械受容チャネルの阻害剤(FM1-43)で消失する.
3.内耳有毛細胞の音感受性の獲得 有毛細胞はいつどのように肝心の音に応じるようになるのであろうか? 有毛細胞の機械受容チャネルが働くと細胞に陽イオンが流れ,その結果耳胞内にはマイクロフォン電位と呼ばれる電場電位が発生する.音刺激に対して最初のマイクロフォン電位が記録されるのは受精後 46 時間であった(図2B)2).ちょうど M 細胞で最初の聴覚応答が記録された時間に一致している.すなわち,内耳有毛細胞から M 細胞までの聴覚回路は受精後 27 時間以前にできていて,しかもその時点で有毛細胞は大きな機械刺激には応答し得るのだが,極めて微弱な音の振動には反応できない.その後20 時間の間に,何かが変わってナノオーダーあるいはサブナノオーダーの振動も受容できるようになり,受精後 46 時間で初めて音に応じられるようになると考えられる.この 20 時間の間に付加される因子が聴覚の獲得にとってキィであるが,その実態はまだ不明である. 我々は2つの可能性を考えている.一つは,微弱な振動に感度のある特殊な機械受容チャネルの発現である.これまでに TRP (transient receptor potential) チャネルの一つである TRPN1 あるいは NompC と名付けられた機械受容チャネルが有力視されていたが,存在位置などに疑問が残っていた.これに対してごく最近,別の膜タンパク質の TMC(Transmembrane channel-like) 1と TMC2 が哺乳動物の内耳の音受容チャネル候補として新たに注目されている 5).
3

これらのチャネルがゼブラフィッシュの内耳有毛細胞の音受容チャネルとして働くかを今後検討しなければならない.もう一つの因子は,耳石の大きさである.発達初期の耳胞内には2つの耳石が存在し,一方は体の平衡を保つのに必要な卵形嚢に位置し,他方は聴覚を担う球形嚢に位置する.将来音の受容に大きな役割を果たす鰾(うきぶくろ)が発達するずっと前の仔魚では,球形嚢の耳石の体積は発達とともに大きくなり,受精後5~6日には卵形嚢の耳石の2~3倍である.大きい耳石ほど慣性が大きく,同じ振動を与えても動きが小さいはずである.軽い有毛細胞が音で動くと重くて動きにくい耳石との間でずれが生じ,その結果繊毛が倒れるであろう.もしそれが正しいならば,本来平衡に働く卵形嚢の耳石を大きくしたら,音に応じるようになるのではないかと考え,球形嚢の耳石を外して卵形嚢の耳石と融合させて大きな卵形嚢の耳石を作ってみた.その結果,驚くべきことに本来からだの傾きを受容する卵形嚢で音応答(マイクロフォン電位)が記録され,数倍の耳石の大きさの違いが音に対する感受性の有無を決めることが示唆された 6).勿論,有毛細胞の機械受容に関連した分子の発現など他の因子が聴覚の獲得にとって決定的に寄与する可能性も十分あるが,聴覚の受容にとっては機械的・力学的な検討が常に必要であろう.
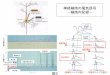
図 3. M 細胞と相同ニューロンの発火特性の違いとその分子基盤.A.後脳に繰り返される相同ニューロン.M 細胞と MiD2/3 は形態学的特徴が同じ相同ニューロンである.スケールバーは 50μm. B.相同ニューロンの発火特性の違い.M 細胞は脱分極入力の開始点で1回だけ発火するのに対して,相同ニューロン (MiD2/3) は脱分極の大きさに依存した周波数で連続的に発火する.M 細胞には2つの低閾値型カリウムチャネルを構成する Kv1.1 と Kv7.4 サブユニットが発現し,MiD2/3 には Kv1.1 のみが発現する.また,Kvβ2 サブユニットは Kv1.1 の発現量を増やす.
4.M 細胞の特異的興奮性の獲得とその分子基盤 内耳で受容された音信号は中枢で特徴抽出されて,必要な出力を生み出す 7).ゼブラフィッシュの音刺激に対する逃避運動を駆動する神経回路では,内耳有毛細胞で受容された音信号は後脳の M 細胞に到達し,M 細胞の発火が反対側の脊髄の運動ニューロンを一斉に興奮させて,逃避運動の最初に起こる胴の屈曲が始まる.聴覚入力は M 細胞以外の後脳網様体脊髄路ニューロン(reticulospinal neurons, RSNs)群(図3A)にも投射し,音刺激に対する逃避運動に寄与すると考えられている.ここで注目すべきは,他の RSNs は脱分極の振幅に応じた頻度で連続的に発火するのに対して,M 細胞は脱分極の開始時にたった 1 発の活動電位しか発生しない(単発発火)という特徴的な性質 8)である(図3B).我々は M 細胞の発火特性が最初は他の RSNs と同様の連続発火型であったのが,聴覚性の逃避運動を獲得した受精後4日目から単発発火型に変化する機構を,電気生理学的および分子生物学的手法で解析した.その結果,低閾値型カリウムチャネル 9) の一つである Kv1.1α サブユニットが M 細胞および M 細胞と形態学的に相同の RSNs に存在し,さらに補助サブユニット 10) Kvβ2 が単発発火特性を示す時期の M 細胞に選択的に現れることを見出した.これらのサブユニットをアフリカツメガエルの卵母細胞に発現させて電気特性を調べたところ,Kv 1.1α サブユニットが膜電位-60 mV 以上で活性化する低閾値型カリウムチャネルを構成し,Kvβ2 との組み合わせによりカリウムの外向き流が顕著に増大することが明らかになった 11).これに加えて,別の低閾値型カリウムチャネルを構成する Kv7.4 も,単発発火する時期の M 細胞に選択的に発現することを見出した.早い時間経過で活性化する Kv1.1 チャネルとは異なって,Kv7.4 チャネルはゆっくりと活性化する特徴も明らかにされた.Kv1.1 と Kv7.4 の両者を阻害すると,M 細胞は他の RSNs と同じように順応のない連続発火型に変化することを見出した.また,Kv1.1 については,単発発火だけでな
4

く,発火までの時間を短縮する効果があり,音刺激からの逃避運動の開始を早くする役割も示唆された.以上の結果から,すばやい逃避運動をトリガーする M 細胞が,大きな脱分極入力に対して短い潜時で単発の活動電位を発生するという特異的な興奮性を獲得するには,これらのサブユニットで構成される2種類の低閾値型カリウムチャネルの発現が必要であり,他の RSNs との主な特性の違いはこの2つで示されることを明らかにした(図3B). 以上のように本研究は,聴覚を獲得するメカニズムについて,モデル脊椎動物のゼブラフィッシュを対象にして,内耳有毛細胞の音受容性の獲得,聴覚回路の形成,中枢ニューロンのの特徴抽出機構の獲得に関して分子生物学的・生理学的・行動学的手法を駆使して解析を進めてきた.ゼブラフィッシュを用いた研究では,これまで他の脊椎動物では困難であった内耳に直接アプローチでき,電気生理学的手法だけでなく遺伝子操作や高度のイメージングやナノマニピュレーションなどを活用することも可能で,長い間謎とされてきた聴覚獲得のメカニズムに関する研究に新しい展開が生まれると期待される.
文 献
1) Kohashi, T. & Oda, Y. : Initiation of Mauthner- or non-Mauthner-mediated fast escape evoked by differentmodes of sensory input. J. Neurosci., 28 : 10641-10653, 2008.
2) Tanimoto, M., Ota, Y., Horikawa, K. & Oda, Y. : Auditory input to CNS is acquired coincidentally withdevelopment of inner ear after formation of functional afferent pathway in zebrafish. J. Neurosci., 29 :2762-2767, 2009.
3) Kohashi, T., Nakata, N. & Oda, Y. : Effective sensory modality activating an escape triggering neuronswitches during early development in zebrafish. J. Neurosci., 32 : 5810-5820, 2012.
4) Tanimoto, M., Ota, Y., Inoue, M. & Oda, Y. : Origin of inner ear hair cells: Morphological and functionaldifferentiation from ciliary cells into hair cells in zebrafish inner ear. J. Neurosci., 31 : 3784-3794, 2011.
5) Pan, B., Géléoc, G. S., Asai, Y., Horwitz, G. C., Kurima, K., Ishikawa, K., Kawashima, Y., Griffith, A. J. & Holt,J. R. : TMC1 and TMC2 are components of the mechanotransduction channel in hair cells of themammalian inner ear. Neuron, 79 : 504-515, 2013.
6) Inoue, M., Tanimoto, M. & Oda, Y. : The role of ear stone size in hair cell acoustic sensory transduction.Sci. Rep., 3 : 2114, 2013.
7) Oertel, D. : The role of timing in the brain stem auditory nuclei of vertebrates. Annu. Rev. Physiol., 61 :497-519, 1999.
8) Nakayama, H. & Oda, Y. : Common sensory inputs and differential excitability of segmentally homologousreticulospinal neurons in the hindbrain. J. Neurosci., 24 : 3199-3209, 2004.
9) Johnston, J., Forsythe, I. D. & Kopp-Scheinpflug, C. : Going native: voltage-gated potassium channelscontrolling neuronal excitability. J. Physiol., 588 : 3187-3200, 2010.
10) Pongs, O. & Schwarz, J. R. : Ancillary subunits associated with voltage-dependent K+ channels. Physiol.Rev., 90 : 755-796, 2010.
11) Watanabe, T., Shimazaki, T., Mishiro, A., Suzuki, T., Hirata, H., Tanimoto, M. & Oda, Y. : Coexpression ofauxiliary Kvß2 subunits with Kv1.1 channels is required for developmental acquisition of unique firingproperties of zebrafish Mauthner cells. J. Neurophysiol., in press.
5