Embed Size (px)
Citation preview

CO R R E S P O N D E N C E
366 VOLUME 11 | NUMBER 4 | APRIL 2005 NATURE MEDICINE
and IFN-γ-induced cell-surface expression of MHC class II molecules after simvastatin treat-ment of various cell types used in our studies. This suggests that an alternative mechanism may contribute to the observed inhibition of MHC class II cell-surface expression by sim-vastatin.
Further suggestions for an alternative mech-anism by which statins show immunomodu-latory properties comes from the reported observations that statins, in addition to their effect on the cell-surface expression of MHC class II molecules, also inhibit expression of immunoregulatory molecules such as adhe-sion molecules, costimulatory molecules and chemokine receptors, diminish the production of proinflammatory cytokines and chemo-kines, and reduce adhesion and proliferation of various cell types5–7. We have made similar observations and have found that statins, in addition to inhibiting expression of MHC class II molecules, also inhibit membrane expres-sion of a number of additional molecules that have important roles in immunity. Subsequent investigations have provided evidence in sup-port of the notion that the effect of statins on cell-surface MHC class II expression is caused by disruption of cholesterol-containing micro-domains (H. Kuipers, P. Biesta, T. Groothuis, J. Neefjes, A. Mommaas & P. van den Elsen, unpublished data). These so-called ‘lipid rafts’ are important for the intracellular transport, recycling and concentration to the cell sur-face of MHC class II molecules9. In general, glycosylphosphatidylinositol-linked proteins are characteristic components of biochemi-cally defined lipid rafts and rely on the integ-rity of these cholesterol-containing vesicles for transport and function at the cell surface. This explains that through disruption of lipid rafts statins may have a much broader effect,
impairing membrane expression and function of a variety of different proteins.
In conclusion, we feel that the immuno-modulatory properties of statins do not seem to be caused through specific inhibition of MHC2TA transcription mediated by the various isotypic promoters. More probably, we hypothesize that disruption of lipid rafts, essential components in a variety of immune processes essential for antigen presentation and subsequent T-cell activation, explains the downregulatory effects of statins on cell-surface expression of MHC class II and other immunoregulatory molecules. This notion is in line with observations made by others10.
ACKNOWLEDGMENT This research is supported by a grant from the Dutch Multiple Sclerosis Research Foundation (grant 00-407MS)
Hedwich F Kuipers1 & Peter J van den Elsen1,2
1Division of Molecular Biology, Department of Immunohematology and Blood Transfusion, Leiden University Medical Center, Leiden, The Netherlands. 2Department of Pathology, VU University Medical Center, Amsterdam, The Netherlands. e-mail: [email protected]
1. Goldstein, J.L. & Brown, M.S. Nature 343, 425 –430 (1990).
2. Vaughan, C.J., Murphy, M.B. & Buckley, B.M. Lancet. 348, 1079–1082 (1996).
3. Mach, F. Mol. Interv. 2, 478–480 (2002). 4. Kwak, B., Mulhaupt, F., Myit, S. & Mach, F. Nat. Med.
6, 1399–1402 (2000).5. Youssef, S. et al. Nature 420, 78–84 (2002).6. Neuhaus, O. et al. Neurology 59, 990–997 (2002).7. Schönbeck, U. & Libby, P. Circulation 109, II-18–II-26
(2004). 8. Reith, W. & Mach, B. Annu. Rev. Immunol. 19, 331–
373 (2001). 9. Poloso, N. & Roche, P. Curr. Opion. Immunol. 16,
103–107 (2004).10. Hillyard, D.Z., Jardine, A.G., McDonald, K.J. &
Cameron, A.J. Atherosclerosis 175, 305–313 (2004)
a b
Figure 2 Simvastatin does not reduce MHC2TA-PIII, MHC2TA-PIV or HLA-DRA promoter activity. (a) U937 monocyte cells transfected with luciferase reporter constructs containing the MHC2TA-PIIIDEL1 (which includes the IFN-γ-inducible upstream region of MHC2TA-PIII), MHC2TA-PIV or HLA-DRA promoters treated with IFN-γ alone or in combination with simvastatin (simva) or simvastatin and L-mevalonate (mev). (b) Raji B cells transfected with luciferase reporter constructs containing the MHC2TA-PIII, MHC2TA-PIV or HLA-DRA promoters, treated with simvastatin alone or in combination with L-mevalonate. Data shown as mean relative light units (RLU) + s.e.m. of one representative of three independent experiments. Asterisks indicate significance of differences according to two-tailed Student t-test: *P < 0.01, **P < 0.05, ***P < 0.1. The pGL3-Basic luciferase reporter plasmid served as a control.
Kwak & Mach reply:A few years ago we reported in Nature Medicine immunomodulatory properties of statins1. We showed that IFN-γ-induced expression of both major histocompatibility complex (MHC) class II and class II transactivator (CIITA) pro-teins was reduced by several statins in primary human endothelial cells and macrophages. In contrast to primary endothelial cells and macrophages, MHC class II is constitutively expressed in, for example, dendritic cells and B cells (Raji). Statins had no effect on consti-tutive MHC class II expression in these cells, making the involvement of MHC2TA promot-ers I and III in statins’ effect unlikely. Promoter IV–specific RNase protection assays and func-tional analysis of promoter IV with a reporter gene assay showed without doubt the involve-ment of this promoter in the statin-induced reduction of inducible MHC class II1,2. Since then, several reports from different research groups confirmed our findings, using mostly primary cells3–5.
The data reported here by Kuipers and van den Elsen seem in apparent contradiction with the aforementioned observations. Kuipers and van den Elsen mention that statins reduced both constitutive and IFN-γ-induced cell-sur-face expression of MHC class II in a variety of different cells. Unfortunately, no information is provided regarding how the statins were prepared (in other words, activated). In con-trast to their findings on cell-surface expres-sion, simvastatin increased the RNAs encoding both MHC class II and CIITA in HeLa and U-937 cells. In addition, reporter gene assays on U-937 cells showed that the statin-induced increase in MHC2TA promoters III and IV and in the MHC class II promoter may be responsi-ble for the increased expression observed at the RNA level. Being non-bone marrow–derived cells, epithelial HeLa cells are not expected to show constitutive MHC class II expression. Consequently, MHC class II RNA (Kuipers’ and van den Elsen’s Fig. 1a) and surface expres-sion (data not shown) observed under basal conditions may result from activation of these cells by infection (viral or mycoplasmic) or other factors, and makes it difficult to ascribe the effect of the statin specifically to one or another mechanism. A mixed response may also be caused by impurity of the cells used. The U-937 cell line is a typical example of such an impure cell line from which subclones with various characteristics can be obtained6,7. The effects observed on CIITA and MHC class II in this cell line (Fig. 1b) may be interpreted as such a mixed response.
The authors propose as a possible explana-tion for their contradictory results between protein surface expression and RNA levels
©20
05 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
urem
edic
ine
CO R R E S P O N D E N C E
NATURE MEDICINE VOLUME 11 | NUMBER 4 | APRIL 2005 367
that statins cause disruption of cholesterol-containing microdomains, this way exerting an additional effect on MHC class II expres-sion. Recently, much attention has been given to lipid rafts as a platform for signal trans-duction. A limited number of papers show data supporting or rejecting the hypothesis that statins disrupt lipid rafts8,9, and we look forward to the paper containing these unpub-lished observations to which the authors refer in this correspondence.
Brenda R Kwak & François Mach
Division of Cardiology, Department of Internal Medicine, University Hospital Geneva, Foundation for Medical Research, 64 Avenue de la Roseraie, Ch-1221 Geneva, Switzerland. E-mail: [email protected]
1. Kwak, B., Mulhaupt, F., Myit, S. & Mach, F. Nat. Med. 6, 1399–1402 (2000).
2. Kwak, B., Mulhaupt, F., Veillard, N., Pelli, G. & Mach, F. Swiss Med Wkly. 131, 41–46 (2001).
3. Sadeghi, M.M. et al. Transplantation 71, 1262–1268 (2001).
4. Youssef, S. et al. Nature 420, 78–84 (2002).5. Neuhaus, O. et al. Neurology 59, 990–997
(2002).6. Willheim, M. et al. Eur. J. Immunol. 25, 3202–3206
(1995).7. Reischl, I.G. et al. Immunol. Lett. 49, 127–131
(1996).8. Hillyard, D.Z., Jardine, A.G., McDonald, K.J. &
Cameron, A.J. Atherosclerosis 172, 219–228 (2004).
9. Ghittoni, R. et al. FASEB J. published online 27 January 2005 (doi:10.1096/fj.04-2702fje).
Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cellsTo the editor:Intramyocardial injection of adult bone marrow–derived stem cells has recently been proposed as a potential therapy to repair damaged myocardium after acute infarction1–
4. The mechanisms underlying their thera-peutic effect have not been clearly defined, with an intense debate over differentiation versus fusion2,5. In this report, we provide evidence for a novel mechanism that involves the secretion of paracrine cytoprotective factor(s) from the stem cells. We have previ-ously reported that rat bone marrow–derived mesenchymal stem cells (MSCs) overexpress-ing the survival gene Akt1 (Akt-MSCs) are
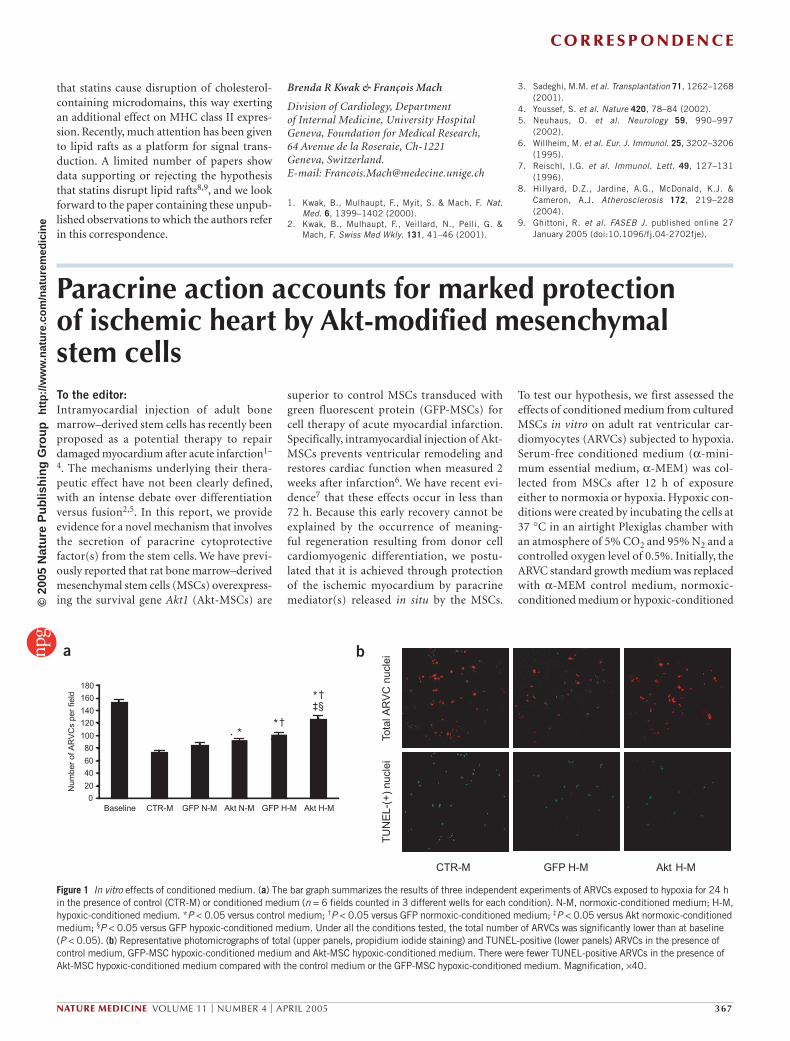
superior to control MSCs transduced with green fluorescent protein (GFP-MSCs) for cell therapy of acute myocardial infarction. Specifically, intramyocardial injection of Akt-MSCs prevents ventricular remodeling and restores cardiac function when measured 2 weeks after infarction6. We have recent evi-dence7 that these effects occur in less than 72 h. Because this early recovery cannot be explained by the occurrence of meaning-ful regeneration resulting from donor cell cardiomyogenic differentiation, we postu-lated that it is achieved through protection of the ischemic myocardium by paracrine mediator(s) released in situ by the MSCs.
To test our hypothesis, we first assessed the effects of conditioned medium from cultured MSCs in vitro on adult rat ventricular car-diomyocytes (ARVCs) subjected to hypoxia. Serum-free conditioned medium (α-mini-mum essential medium, α-MEM) was col-lected from MSCs after 12 h of exposure either to normoxia or hypoxia. Hypoxic con-ditions were created by incubating the cells at 37 °C in an airtight Plexiglas chamber with an atmosphere of 5% CO2 and 95% N2 and a controlled oxygen level of 0.5%. Initially, the ARVC standard growth medium was replaced with α-MEM control medium, normoxic-conditioned medium or hypoxic-conditioned
Figure 1 In vitro effects of conditioned medium. (a) The bar graph summarizes the results of three independent experiments of ARVCs exposed to hypoxia for 24 h in the presence of control (CTR-M) or conditioned medium (n = 6 fields counted in 3 different wells for each condition). N-M, normoxic-conditioned medium; H-M, hypoxic-conditioned medium. *P < 0.05 versus control medium; †P < 0.05 versus GFP normoxic-conditioned medium; ‡P < 0.05 versus Akt normoxic-conditioned medium; §P < 0.05 versus GFP hypoxic-conditioned medium. Under all the conditions tested, the total number of ARVCs was significantly lower than at baseline (P < 0.05). (b) Representative photomicrographs of total (upper panels, propidium iodide staining) and TUNEL-positive (lower panels) ARVCs in the presence of control medium, GFP-MSC hypoxic-conditioned medium and Akt-MSC hypoxic-conditioned medium. There were fewer TUNEL-positive ARVCs in the presence of Akt-MSC hypoxic-conditioned medium compared with the control medium or the GFP-MSC hypoxic-conditioned medium. Magnification, ×40.
Baseline CTR-M GFP N-M Akt N-M GFP H-M Akt H-M0
20
40
60
80
100
120
140
160
180
*
*†‡§
*†
Num
ber
of A
RV
Cs
per
field
CTR-M GFP H-M Akt H-M
Tota
l AR
VC
nuc
lei
TU
NE
L-(+
) nu
clei
a b
©20
05 N
atur
e P
ublis
hing
Gro
up
http
://w
ww
.nat
ure.
com
/nat
urem
edic
ine